
Abstract
Measures of tree balance play an important role in the analysis of phylogenetic trees. One of the oldest and most popular indices in this regard is the Colless index for rooted bifurcating trees, introduced by Colless (Syst Zool 31:100–104, 1982). While many of its statistical properties under different probabilistic models for phylogenetic trees have already been established, little is known about its minimum value and the trees that achieve it. In this manuscript, we fill this gap in the literature. To begin with, we derive both recursive and closed expressions for the minimum Colless index of a tree with n leaves. Surprisingly, these expressions show a connection between the minimum Colless index and the so-called Blancmange curve, a fractal curve. We then fully characterize the tree shapes that achieve this minimum value and we introduce both an algorithm to generate them and a recurrence to count them. After focusing on two extremal classes of trees with minimum Colless index (the maximally balanced trees and the greedy from the bottom trees), we conclude by showing that all trees with minimum Colless index also have minimum Sackin index, another popular balance index.








Similar content being viewed by others
Notes
We adopt the convention that 0 belongs to the set \({\mathbb {N}}\) of natural numbers, and, for any given \(m\in {\mathbb {N}}{\setminus }\{0\}\), we use the notation \({\mathbb {N}}_{\geqslant m}:=\{n\in {\mathbb {N}}\mid n\geqslant m\}\).
References
Agapow P, Purvis A (2002) Power of eight tree shape statistics to detect nonrandom diversification: a comparison by simulation of two models of cladogenesis. Syst Biol 51:866–872
Aldous D (1996) Probability distributions on cladograms. In: Aldous D, Pemantle R (eds) Random discrete structures. The IMA volumes in mathematics and its applications, vol 76. Springer, New York, pp 1–18
Aldous D (2001) Stochastic models and descriptive statistics for phylogenetic trees, from Yule to today. Stat Sci 16:23–34
Allaart PC, Kawamura K (2012) The Takagi function: a survey. Real Anal Exchange 37:1–54
Avino M, Garway TN, et al (2018) Tree shape-based approaches for the comparative study of cophylogeny. bioRxiv 10.1101/388116
Blum MG, François O (2005) On statistical tests of phylogenetic tree imbalance: the Sackin and other indices revisited. Math Biosci 195:141–153
Blum MG, François O (2006) Which random processes describe the tree of life? a large-scale study of phylogenetic tree imbalance. Syst Biol 55:685–691
Blum MGB, François O, Janson S (2006) The mean, variance and limiting distribution of two statistics sensitive to phylogenetic tree balance. Ann Appl Probab 16:2195–2214
Bortolussi N, Durand E, Blum M, François O (2005) apTreeshape: statistical analysis of phylogenetic tree shape. Bioinformatics 22:363–364
Brower AVZ, Rindal E (2013) Reality check: a reply to Smith. Cladistics 29:464–465
Cardona G, Mir A, Rosselló F (2013) Exact formulas for the variance of several balance indices under the Yule model. J Math Biol 67:1833–1846
Chalmandrier L, Albouy C et al (2018) Comparing spatial diversification and meta-population models in the Indo-Australian Archipelago. R Soc Open Sci 5:171366
Colless D (1982) Review of phylogenetics: the theory and practice of phylogenetic systematics. Syst Zool 31:100–104
Colless D (1995) Relative symmetry of cladograms and phenograms: an experimental study. Syst Biol 44:102–108
Coronado TM, Mir A, Rosselló F, Valiente G (2019) A balance index for phylogenetic trees based on rooted quartets. J Math Biol 79:1105–1148
Cunha T, Giribet G (2019) A congruent topology for deep gastropod relationships. Proc R Soc B 286:20182776
Drummond AJ, Ho SYW, Phillips MJ, Rambaut A (2006) Relaxed phylogenetics and dating with confidence. PLoS Biol 4:e88
Duchene S, Bouckaert R, Duchene DA, Stadler T, Drummond AJ (2018) Phylodynamic model adequacy using posterior predictive simulations. Syst Biol 68:358–364
Farris J, Källersjö M (1998) Asymmetry and explanations. Cladistics 14:159–166
Felsenstein J (2004) Inferring phylogenies. Sinauer Associates Inc, Sinauer
Fischer M (2018) Extremal values of the Sackin balance index for rooted binary trees. arXiv preprint arXiv:1801.10418v3
Fischer M, Liebscher V (2015) On the balance of unrooted trees. arXiv preprint arXiv:1510.07882
Ford DJ (2005) Probabilities on cladograms: introduction to the alpha model. PhD thesis, Stanford University. arXiv preprint arXiv:math/0511246
Fusco G, Cronk QC (1995) A new method for evaluating the shape of large phylogenies. J Theor Biol 175:235–243
Futuyma DJ (ed) (1999) Evolution, science and society: evolutionary biology and the national research agenda. The State University of New Jersey, New Jersey
Goloboff PA, Arias JS, Szumik CA (2017) Comparing tree shapes: beyond symmetry. Zool Scr 46:637–648
Hayati M, Shadgar B, Chindelevitch L (2019) A new resolution function to evaluate tree shape statistics. PloS ONE 14:e0224197
Heard SB (1992) Patterns in tree balance among cladistic, phenetic, and randomly generated phylogenetic trees. Evolution 46:1818–1826
Hillis D, Bull J, White M et al (1992) Experimental phylogenetics: generation of a known phylogeny. Science 255:589–592
Holton T, Wilkinson M, Pisani D (2014) The shape of modern tree reconstruction methods. Syst Biol 63:436–441
Kayondo H, Mwalili S, Mango J (2019) Inferring multi-type birth-death parameters for a structured host population with application to HIV epidemic in Africa. Comput Mol Biosci 9:108–131
Kingman JFC (1982) The coalescent. Stochastic Process Appl 13:235–248
Kirkpatrick M, Slatkin M (1993) Searching for evolutionary patterns in the shape of a phylogenetic tree. Evolution 47:1171–1181
Kubo T, Iwasa Y (1995) Inferring the rates of branching and extinction from molecular phylogenies. Evolution 49:694–704
Matsen F (2006) A geometric approach to tree shape statistics. Syst Biol 55:652–61
McKenzie A, Steel M (2000) Distributions of cherries for two models of trees. Math Biosci 164:81–92
Metzig C, Ratmann O, Bezemer D, Colijn C (2019) Phylogenies from dynamic networks. PLoS Comput Biol 15:e1006761
Mir A, Roselló F, Rotger L (2013) A new balance index for phylogenetic trees. Math Biosci 241:125–136
Mir A, Rotger L, Rosselló F (2018) Sound Colless-like balance indices for multifurcating trees. PLoS ONE 13:e0203401
Mooers AO, Heard SB (1997) Inferring evolutionary process from phylogenetic tree shape. Q Rev Biol 72:31–54
Nelson MI, Holmes EC (2007) The evolution of epidemic influenza. Nat Rev Genetics 8:196–205
Piel WH, Chan L, Dominus MJ et al (2009) TreeBASE v.2: a database of phylogenetic knowledge. In: e-BioSphere 2009
Poon AF (2015) Phylodynamic inference with kernel ABC and its application to HIV epidemiology. Mol Biol Evol 32:2483–2495
Purvis A (1996) Using interspecies phylogenies to test macroevolutionary hypotheses. In: Harvey PH, Brown AJL, Maynard Smith J, Nee S (eds) New uses for new phylogenies. Oxford University Press, Oxford, pp 153–168
Purvis A, Fritz S, Rodríguez J, Harvey P, Grenyer R (2011) The shape of mammalian phylogeny: patterns, processes and scales. Philos Trans R Soc B 366:2462–2477
Purvis A, Katzourakis A, Agapow P-M (2002) Evaluating phylogenetic tree shape: two modifications to Fusco & Cronk’s method. J Theor Biol 214:99–103
Rindal E, Brower AVZ (2011) Do model-based phylogenetic analyses perform better than parsimony? A test with empirical data. Cladistics 27:331–334
Rogers JS (1993) Response of Colless’s tree imbalance to number of terminal taxa. Syst Biol 42:102
Sackin MJ (1972) “Good” and “bad” phenograms. Syst Zool 21:225–226
Saulnier E, Alizon S, Gascuel O (2016) Assessing the accuracy of approximate bayesian computation approaches to infer epidemiological parameters from phylogenies. bioRxiv, 050211. https://doi.org/10.1101/050211
Savage HM (1983) The shape of evolution: systematic tree topology. Biol J Linn Soc 20:225–244
Shao K, Sokal R (1990) Tree balance. Syst Zool 39:266–276
Semple C, Steel M (2003) Phylogenetics. Oxford University Press, Oxford
Sloane NJA (1964) The on-line encyclopedia of integer sequences (OEIS). http://oeis.org. Last accessed 8 July 2019
Slowinski J (1990) Probabilities of \(n\)-trees under two models: a demonstration that asymmetrical interior nodes are not improbable. Syst Zool 39:89–94
Sober E (1993) Experimental tests of phylogenetic inference methods. Syst Biol 42:85–89
Stam E (2002) Does imbalance in phylogenies reflect only bias? Evolution 56:1292–1295
Steel M (2016) Phylogeny: discrete and random processes in evolution. SIAM, New York
Stich M, Manrubia SC (2009) Topological properties of phylogenetic trees in evolutionary models. Eur Phys J B 70:583–592
Takagi T (1901) A simple example of continuous function without derivative. Tokyo Sugaku Butsurigakkwai Hokoku 1:F176–F177
Verboom G, Boucher F, Ackerly D et al (2019) Species selection regime and phylogenetic tree shape. Syst Biol (in press). https://doi.org/10.1093/sysbio/syz076
Vos RA, Balhoff JP, Caravas JA et al (2012) NeXML: rich, extensible, and verifiable representation of comparative data and metadata. Syst Biol 61:675–689
Willis JC, Yule GU (1922) Some statistics of evolution and geographical distribution in plants and animals, and their significance. Nature 109:177–179
Wu T, Choi K (2015) On joint subtree distributions under two evolutionary models. Theor Popul Biol 108:13–23
Acknowledgements
Tomás M. Coronado and Francesc Rosselló thank the Spanish Ministry of Science, Innovation and Universities, the Spanish Research Agency, and the European Regional Development Fund through Projects DPI2015-67082-P and PGC2018-096956-B-C43 (FEDER/MICINN/AEI). Moreover, Mareike Fischer thanks the joint research project DIG-IT! supported by the European Social Fund (ESF), reference: ESF/14-BM-A55-0017/19, and the Ministry of Education, Science and Culture of Mecklenburg-Vorpommern, Germany. Additionally, Lina Herbst thanks the state Mecklenburg-Western Pomerania for a Landesgraduierten-Studentship and Kristina Wicke thanks the German Academic Scholarship Foundation for a studentship. Moreover, we thank the anonymous reviewers and the editors for their helpful comments on an earlier version of this manuscript.
Author information
Authors and Affiliations
Corresponding author
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Appendices
Appendices
1.1 A.1 Proof of Proposition 2
Recall that, for every \(n \in {\mathbb {N}}_{\geqslant 2}\),
We shall establish the following result.
Proposition 2
For every \(n\geqslant 2\) and for every \(n_a,n_b\in {\mathbb {N}}_{\geqslant 1}\) such that \(n_a\geqslant n_b\) and \(n_a+n_b=n\):
- (1)
If \(n_a=n_b=n/2\), then \((n_a,n_b)\in QB(n)\) always.
- (2)
If \(n_a>n_b\), then \((n_a,n_b)\in QB(n)\) if, and only if, one of the following three conditions is satisfied:
There exist \(k\in {\mathbb {N}}\) and \(p\in {\mathbb {N}}_{\geqslant 1}\) such that \(n=2^k(2p+1)\), \(n_a=2^k(p+1)\) and \(n_b=2^kp\).
There exist \(k\in {\mathbb {N}}\), \(l\in {\mathbb {N}}_{\geqslant 2}\), \(p\in {\mathbb {N}}_{\geqslant 1}\), and \(t\in {\mathbb {N}}\), \(0\leqslant t<2^{l-2}\), such that \(n=2^k(2^l(2p+1)+2t+1)\), \(n_a=2^{k+l}(p+1)\), and \(n_b=2^{k}(2^lp+2t+1)\).
There exist \(k\in {\mathbb {N}}\), \(l\in {\mathbb {N}}_{\geqslant 2}\), \(p\in {\mathbb {N}}_{\geqslant 1}\), and \(t\in {\mathbb {N}}\), \(0\leqslant t<2^{l-2}\), such that \(n=2^k(2^l(2p+1)-(2t+1))\), \(n_a=2^k(2^l(p+1)-(2t+1))\), and \(n_b=2^{k+l}p\).
The proof of this proposition relies on several auxiliary lemmas. In order to simplify the language in their statements and proofs, throughout this section we systematically assume, without any further notice, that the symbols j, k, m, n, p, s, t, and x, possibly with subscripts or superscripts, always represent natural numbers.
Lemma 7
Let \(s=2^ks_0\) with \(k\geqslant 1\) and \(s_0\geqslant 1\). Then, for every \(m\geqslant 1\), \((m+s,m)\in QB(2m+s)\) if, and only if, \(m=2^km_0\), for some \(m_0\geqslant 1\) such that \((m_0+s_0,m_0)\in QB(2m_0+s_0)\).
Proof
We prove the equivalence in the statement by induction on the exponent \(k\geqslant 1\). Recall that, by Remark 2.(b), if \(s\geqslant 1\) is even and \(c_{m+s}+c_m+s=c_{2m+s}\), then m must be even, too. Therefore, if \(s=2t_0\), then \(m=2m_1\) for some \(m_1\geqslant 1\), and then, since
and \(c_{4m_1+2t_0}=2c_{2m_1+t_0}\), the equality \(c_{m+s}+c_m+s=c_{2m+s}\) is equivalent to the equality \(c_{m_1+t_0}+c_{m_1}+t_0=c_{2m_1+t_0}\). This proves the equivalence in the statement when \(k=1\).
Now, assume that this equivalence is true for the exponent \(k-1\), and let \(s=2^ks_0\). Then, by the case \(k=1\), \(c_{m+s}+c_m+s=c_{2m+s}\) if, and only if, \(m=2m_1\) for some \(m_1\geqslant 1\) such that
and, by the induction hypothesis, this last equality holds if, and only if, \(m_1=2^{k-1}m_0\) for some \(m_0\geqslant 1\) such that \(c_{m_0+s_0}+c_{m_0}+s_0=c_{2m_0+s_0}\). Combining both equivalences we obtain the equivalence in the statement, thus proving the inductive step. \(\square \)
Lemma 8
Let \(s=2^{j+1}-(2t+1)\) be an odd integer, with \(j={\lfloor }\log _2(s){\rfloor }\) and \(0\leqslant t<2^{j-1}\). Then, for every \(m\geqslant 1\), \((2m+s,2m)\in QB(4m+s)\) if, and only if, \(m=2^{j}p\) for some \(p\geqslant 1\).
Proof
We prove the equivalence in the statement by induction on s. When \(s=1=2^{1}-1\), so that \(j=t=0\), the equivalence says that
for every \(m\geqslant 1\), which is true by Corollary 3.
Assume now that the equivalence is true for every odd natural number \(s'<s\) and for every m, and let us prove it for \(s=2^{j+1}-(2t+1)\) with \(0\leqslant t<2^{j-1}\). We have that
and since, by Eq. (2), \(c_{m+2^j-t}+c_m+2^j-t\geqslant c_{2m+2^j-t}\) and \(c_{m+2^j-t-1}+c_m+2^j-t-1\geqslant c_{2m+2^j-t-1}\), we have that \(c_{2m+s}+c_{2m}+s=c_{4m+s}\) if, and only if, the following two identities are satisfied:
So, we must prove that Eqns. (13) and (14) hold if, and only if, \(m=2^{j}p\) for some \(p\geqslant 1\). We distinguish two subcases, depending on the parity of t:
If \(t=2x\) for some \(0\leqslant x<2^{j-2}\), then Eq. (13) and Lemma 7 imply that m is even, say \(m=2m_0\), and then (14) says
$$\begin{aligned} c_{2m_0+2^j-2x-1}+c_{2m_0}+2^j-2x-1= c_{4m_0+2^j-2x-1}, \end{aligned}$$(15)which, by induction, is equivalent to \(m_0=2^{j-1}p\) for some \(p\geqslant 1\), i.e. to \(m=2^{j}p\) for some \(p\geqslant 1\). So, to complete the proof of the desired equivalence, it remains to prove that if \(m=2^{j}p\), then Eq. (13) holds. If \(t=0\), this equality says
$$\begin{aligned} c_{2^{j}p+2^j}+c_{2^{j}p}+2^j=c_{2^{j+1}p+2^j} \end{aligned}$$and it is a direct consequence of Lemma 7 and Corollary 3. So, assume that \(t>0\) and write it as \(t=2^i(2x_0+1)\) with \(1\leqslant i<j-1\) and \(x_0<2^{j-i-2}\). Then
$$\begin{aligned}&c_{m+2^j-t}+c_m+2^j-t \\&\quad = c_{2^{j}p+2^j-2^i(2x_0+1)} +c_{2^{j}p}+2^j-2^i(2x_0+1)\\&\quad = 2^i\big (c_{2^{j-i}p+2^{j-i}-2x_0-1}+c_{2^{j-i}p}+2^{j-i}-2x_0-1\big )\\&\quad = 2^ic_{2^{j-i+1}p+2^{j-i}-2x_0-1} \text{(by } \text{ the } \text{ induction } \text{ hypothesis) }\\&\quad = c_{2^{j+1}p+2^{j}-2^i(2x_0+1)}=c_{2m+2^j-t}. \end{aligned}$$If \(t=2x+1\) for some \(0\leqslant x< 2^{j-2}\), then Eq. (14) and Lemma 7 imply that m is even, say \(m=2m_0\), and then it is Eq. (13) which becomes Eq. (15) above, which, in turn, by induction is equivalent to \(m_0=2^{j-1}p\) for some \(p\geqslant 1\), that is, to \(m=2^{j}p\) for some \(p\geqslant 1\). Thus, to complete the proof of the desired equivalence, it remains to prove that if \(m=2^{j}p\), then (14) holds. Now:
$$\begin{aligned}&c_{m+2^j-t-1}+c_m+2^j-t-1\\&\quad = c_{2^{j}p+2^j-2x-2}+c_{2^{j}p}+2^j-2x-2\\&\quad =2\big (c_{2^{j-1}p+2^{j-1}-x-1}+c_{2^{j-1}p}+2^{j-1}-x-1\big ) \end{aligned}$$If x is even, say \(x=2x_0\), then, since \(x_0<2^{j-3}\), the induction hypothesis implies that
$$\begin{aligned}&2\big (c_{2^{j-1}p+2^{j-1}-x-1}+c_{2^{j-1}p}+2^{j-1}-x-1\big )\\&\quad =2 c_{2^{j}p+2^{j-1}-x-1} = c_{2^{j+1}p+2^{j}-2x-2}= c_{2m+2^j-t-1}. \end{aligned}$$And if x is odd, write it as \(x=2^i(2t_0+1)-1\) for some \(1\leqslant i<j-1\) (and notice that \(x<2^{j-2}\) implies \(t_0<2^{j-i-3}\)) and then
$$\begin{aligned}&2\big (c_{2^{j-1}p+2^{j-1}-x-1}+c_{2^{j-1}p}+2^{j-1}-x-1\big )\\&\quad = 2\big (c_{2^{j-1}p+2^{j-1}-2^i(2t_0+1)}+c_{2^{j-1}p}+2^{j-1}-2^i(2t_0+1)\big )\\&\quad =2\cdot 2^i\big (c_{2^{j-i-1}p+2^{j-i-1}-(2t_0+1)}+c_{2^{j-i-1}p}+2^{j-i-1}-(2t_0+1)\big )\\&\quad = 2^{i+1} c_{2^{j-i}p+2^{j-i-1}-(2t_0+1)} \text{(by } \text{ the } \text{ induction } \text{ hypothesis) }\\&\quad = c_{2^{j+1}p+2^{j}-2^{i+1}(2t_0+1)}=c_{2^{j+1}p+2^{j}-2x-2}\\&\quad =c_{2m+2^j-t-1} \end{aligned}$$This completes the proof of the desired equivalence when t is odd.
So, the inductive step is true in all cases. \(\square \)
Lemma 9
Let \(s=2^{j+1}-(2t+1)\) be an odd integer, with \(j={\lfloor }\log _2(s){\rfloor }\) and \(0\leqslant t<2^{j-1}\). Then, for every \(m\geqslant 0\), \((2m+1+s,2m+1)\in QB(4m+2+s)\) if, and only if, either \(m=2^{j}p+t\) for some \(p\geqslant 1\) or \(s=1\) (i.e. \(j=t=0\)) and \(m=0\).
Proof
We also prove the equivalence in this statement by induction on s. When \(s=1=2^{1}-1\), the equivalence says that \(c_{2m+2}+c_{2m+1}+1=c_{4m+3}\) for every \(m\geqslant 0\), which is true by Corollary 3.
Assume now that the equivalence is true for every odd natural number \(1\leqslant s'<s\) and for every \(m\geqslant 0\), and let us prove it for \(s=2^{j+1}-(2t+1)\geqslant 3\) with \(0\leqslant t<2^{j-1}\). In this case, m cannot be 0, because, by Remark 2.(a), \((s+1,1)\in QB(s+2)\) if, and only if, \(s=1\). So, we can consider only the case \(m\geqslant 1\). Then, we have that
and since, by Eq. (2), \(c_{m+2^j-t}+c_m+2^j-t\geqslant c_{2m+2^j-t}\) and \(c_{m+2^j-t}+c_{m+1}+2^j-t-1\geqslant c_{2m+2^j-t+1}\), we have that \(c_{2m+1+s}+c_{2m+1}+s=c_{4m+2+s}\) if, and only if,
So, we must prove that Eqns. (16) and (17) hold for \(m\geqslant 1\) if, and only if, \(m=2^{j}p+t\) for some \(p\geqslant 1\). We distinguish again two subcases, depending on the parity of t:
If \(t=2x\) for some \(0\leqslant x<2^{j-2}\), then Eq. (16) and Lemma 7 imply that m is even, say \(m=2m_0\) with \(m_0\geqslant 1\), and then Eq. (17) can be written
$$\begin{aligned} c_{2m_0+1+2^j-2x-1}+c_{2m_0+1}+2^j-2x-1= c_{4m_0+2+2^j-2x-1} \end{aligned}$$which, by induction, is equivalent to \(m_0=2^{j-1}p+x\) for some \(p\geqslant 1\), that is, to \(m=2m_0=2^{j}p+t\) for some \(p\geqslant 1\). Hence, to complete the proof of the desired equivalence, it remains to check that if \(m=2^{j}p+t\), then Eq. (16) holds. Now, if \(x=0\), so that \(m=2^{j}p\), Corollary 3 and Lemma 7 clearly imply Eq. (16) (cf. the case when t is even in the proof of Lemma 8). So, assume that \(x>0\) and write it as \(x=2^i(2y_0+1)\) with \(0\leqslant i< j-2\) and \(y_0<2^{j-i-3}\). Then
$$\begin{aligned}&c_{m+2^j-t}+c_m+2^j-t\\&\quad =c_{2^{j}p+2x+2^j-2x}+c_{2^{j}p+2x}+2^j-2x\\&\quad = c_{2^{j}p+2^{i+1}(2y_0+1)+2^{j}-2^{i+1}(2y_0+1)}\\&\qquad +c_{2^{j}p+2^{i+1}(2y_0+1)}+2^{j}-2^{i+1}(2y_0+1)\\&\quad =2^{i+1}\big (c_{2^{j-i-1}p+2y_0+1+2^{j-i-1} -(2y_0+1)}\\&\qquad +c_{2^{j-i-1}p+2y_0+1}+2^{j-i-1}-(2y_0+1)\big )\\&\quad =2^{i+1} c_{2^{j-i}p+4y_0+2+2^{j-i-1}-(2y_0+1)}\quad \text{(by } \text{ the } \text{ induction } \text{ hypothesis) }\\&\quad =c_{2^{j+1}p+2^{j}+2^{i+1}(2y_0+1)}=c_{2^{j+1}p+2^{j}+2x}\\&\quad =c_{2m+2^j-t} \end{aligned}$$as we wanted to prove.
If \(t=2x+1\) for some \(0\leqslant x<2^{j-2}\), Eq. (17) and Lemma 7 imply that \(m+1\) is even, and then m is odd, say \(m=2m_0+1\) for some \(m_0\geqslant 0\), and Eq. (16) can be written
$$\begin{aligned} c_{2m_0+1+2^j-2x-1}+c_{2m_0+1}+2^j-2x-1= c_{4m_0+2+2^j-2x-1}. \end{aligned}$$(18)Now, if \(m_0=0\), Remark 2.(a) implies that this equality holds if, and only if, \(2^j-2x-1=1\) which, under the condition \(0\leqslant x<2^{j-2}\), only happens when \(j=1\) and \(x=0\), but then \(t=1=2^{j-1}\) against the assumption that \(t<2^{j-1}\). Therefore \(m_0\) must be at least 1. Then, by induction, Identity (18) is equivalent to \(m_0=2^{j-1}p+x\) for some \(p\geqslant 1\), that is, to \(m=2m_0+1=2^{j}p+2x+1=2^{j}p+t\) for some \(p\geqslant 1\). So, to complete the proof of the desired equivalence, it remains to check that if \(m=2^{j}p+t\), then Eq. (17) holds. Now, in the current situation:
$$\begin{aligned}&c_{m+2^j-t}+c_{m+1}+2^j-t-1\\&\quad =c_{2^{j}p+2x+1+2^j-2x-1}+c_{2^{j}p+2x+2}+2^j-2x-2\\&\quad =c_{2^{j}p+2^j}+c_{2^{j}p+2x+2}+2^j-2x-2\\&\quad = 2\big (c_{2^{j-1}p +2^{j-1}}+c_{2^{j-1}p+x+1}+2^{j-1}-x-1\big )\\&\quad = 2\big (c_{(2^{j-1}p+x+1)+(2^{j-1}-x-1)}+c_{2^{j-1}p+x+1}+2^{j-1}-x-1\big )=(**) \end{aligned}$$If x is even, say \(x=2x_0\) with \(0\leqslant x_0<2^{j-3}\), then
$$\begin{aligned} (**)&=2\big (c_{(2^{j-1}p+2x_0+1)+(2^{j-1}-2x_0-1)}+c_{2^{j-1}p+2x_0+1}+2^{j-1}-2x_0-1\big )\\&= 2c_{2^{j}p+2(2x_0+1)+2^{j-1}-(2x_0+1)} \quad \text{(by } \text{ the } \text{ induction } \text{ hypothesis) }\\&=c_{2^{j+1}p+2^{j}+4x_0+2} =c_{2m+2^j-t+1}. \end{aligned}$$And if x is odd, write it as \(x=2^i(2t_0+1)-1\) with \(1\leqslant i<j-1\) and \(t_0<2^{j-i-3}\), and then
$$\begin{aligned} (**)&=2\big (c_{2^{j-1}p+2^i(2t_0+1)+2^{j-1}-2^i(2t_0+1)}\\&\qquad +c_{2^{j-1}p+2^i(2t_0+1)}+2^{j-1}-2^i(2t_0+1)\big )\\&= 2^{i+1}\big (c_{2^{j-i-1}p+2t_0+1+2^{j-i-1}-(2t_0+1)}\\&\qquad +c_{2^{j-i-1}p+2t_0+1}+2^{j-i-1}-(2t_0+1)\big )\\&= 2^{i+1}c_{2^{j-i}p+4t_0+2+2^{j-i-1}-(2t_0+1)} \quad \text{(by } \text{ the } \text{ induction } \text{ hypothesis) }\\&= c_{2^{j+1}p+2^{i+1}(2t_0+1)+2^{j}}=c_{2^{j+1}p+2x+2+2^{j}}\\&=c_{2m+2^j-t+1} \end{aligned}$$This completes the proof of the desired equivalence when t is odd.
\(\square \)
We are now in a position to proceed with the proof of Proposition 2. Assertion (1) in it is a direct consequence of Corollary 3. So, assume \(n_a>n_b\) and set \(s=n_a-n_b\), so that \(n_a=n_b+s\). Then:
- (a)
If \(s=1\), then, by Lemma 9, \(c_{n_a}+c_{n_b}+n_a-n_b=c_{n_a+n_b}\) for every \(n_b\geqslant 1\).
- (b)
If \(s>1\) is odd, write it as \(s=2^{j+1}-(2t+1)\), with \(j={\lfloor }\log _2(s){\rfloor }\geqslant 1\) and \(0\leqslant t<2^{j-1}\). Then, by Lemmas 8 and 9, \(c_{n_a}+c_{n_b}+n_a-n_b=c_{n_a+n_b}\) if, and only if, either \(n_b=2^{j+1}p\) or \(n_b=2^{j+1}p+2t+1\), for some \(p\geqslant 1\).
- (c)
If \(s\geqslant 2\) is even, write it as \(s=2^ks_0\), with \(k\geqslant 1\) the largest exponent of a power of 2 that divides s and \(s_0\) an odd integer, and write the latter as \(s_0=2^{j+1}-(2t+1)\) with \(j={\lfloor }\log _2(s_0){\rfloor }\geqslant 0\) and \(0\leqslant t<2^{j-1}\). Then, by Lemma 7, \(c_{n_a}+c_{n_b}+n_a-n_b=c_{n_a+n_b}\) if, and only if, \(n_b=2^km\), for some \(m\geqslant 1\) such that \(c_{m+s_0}+c_m+s_0=c_{2m+s_0},\) and then:
If \(s_0=1\) (equivalently, if \(j=0\)), \(c_{m+s_0}+c_m+s_0=c_{2m+s_0}\) for every \(m\geqslant 1\) and therefore, in this case, \(c_{n_a}+c_{n_b}+n_a-n_b=c_{n_a+n_b}\) for every \(n_b=2^km\) with \(m\geqslant 1\).
If \(s_0>1\) (equivalently, if \(j>0\)), Lemmas 8 and 9 imply that \(c_{m+s_0}+c_m+s_0=c_{2m+s_0}\) if, and only if, \(m=2^{j+1}p\) or \(m=2^{j+1}p+2t+1\), for some \(p\geqslant 1\). Therefore, in this case, \(c_{n_a}+c_{n_b}+n_a-n_b=c_{n_a+n_b}\) if, and only if, \(n_b=2^{k+j+1}p\) or \(n_b=2^k(2^{j+1}p+2t+1)\), for some \(p\geqslant 1\).
Combining the three cases, and taking \(k=0\) in the odd s case, we conclude that
if, and only if, writing \(n_a-n_b=2^k(2^{j+1}-(2t+1))\) (for some \(k\geqslant 0\), \(j\geqslant 0\), and \(0\leqslant t<2^{j-1}\)),
If \(j=0\), then \(n_b=2^kp\) for some \(p\geqslant 1\), in which case \(n_a=2^k(p+1)\) and \(n=2^{k}(2p+1)\).
If \(j>0\), then there exists some \(p\geqslant 1\) for which one of the following conditions holds:
\(n_b=2^{k+j+1}p\), in which case \(n_a=2^k(2^{j+1}(p+1)-(2t+1))\) and \(n=2^k(2^{j+1}(2p+1)-(2t+1))\).
\(n_b=2^k(2^{j+1}p+2t+1)\), \(n_a=2^{k+j+1}(p+1)\) and \(n=2^k(2^{j+1}(2p+1)+ 2t+1)\).
This is equivalent to the expressions for \(n_a\) and \(n_b\) in option (2) in the statement (replacing \(j+1\) with \(j>0\) by \(l\geqslant 2\)).
This completes the proof of Proposition 2.
1.2 A.2 Proof of Proposition 5
This appendix is devoted to establish the following result.
Proposition 5
Let \(T_n^{ gfb }=(T_a,T_b)\) be a GFB tree with \(n\geqslant 2\), \(T_a\in {\mathcal {T}}_{n_a}\), \(T_b\in {\mathcal {T}}_{n_b}\) and \(n_a\geqslant n_b\). Let \(n=2^m+p\) with \(m=\lfloor \log _2(n)\rfloor \) and \(0\leqslant p<2^m\). Then, we have:
- (i)
If \(0\leqslant p\leqslant 2^{m-1}\), then \(n_a = 2^{m-1}+p\), \(n_b = 2^{m-1}\) and \(T_b\) is fully symmetric.
- (ii)
If \(2^{m-1}\leqslant p<2^m\), \(n_a = 2^{m}\), \(n_b=p\) and \(T_a\) is fully symmetric.
The proof of this proposition requires of the following lemma. The idea guiding its proof is illustrated in Fig. 9.
Lemma 10
Let \(n\geqslant 3\) be an odd natural number. Then, \(T_n^{ gfb }\) shares a maximal pending subtree with \(T_{n-1}^{ gfb }\) and a maximal pending subtree with \(T_{n+1}^{ gfb }\).
Proof
Since \(n\geqslant 3\) is odd, the first \((n-1)/{2}\) iterations of the loop in Algorithm 2 result in \({(n-1)}/{2}\) cherries and a single node, which in the \((n+1)/{2}\)-th iteration is clustered with a cherry to form a tree with 3 leaves. From this moment on, as the algorithm continues clustering trees, in each i-th iteration there will be one, and only one, tree \(T_i^{odd}\) with an odd number s(i) of leaves. Note now that, on the one hand, this unique tree with s(i) leaves is treated by the algorithm like a tree with \(s(i)-1\) leaves, except that it is clustered as late as possible, i.e. when all other trees in \( treeset \) with \(s(i)-1\) leaves (if there are any) have already been clustered. On the other hand, however, this tree is also treated by the algorithm like a tree with \(s(i)+1\) leaves, except that it is clustered as early as possible, i.e. before any other elements in \( treeset \) with \(s(i)+1\) leaves (if there are any) get clustered. So, to summarize, after the first \(i\geqslant (n+1)/{2}\) iterations of the loop, \( treeset \) contains a unique tree \(T_i^{odd}\) with an odd number s(i) of leaves, which at the same time
- (i)
is treated like a tree with \(s(i)-1\) leaves, but is clustered as late as possible;
- (ii)
is treated like a tree with \(s(i)+1\) leaves, but is clustered as soon as possible.

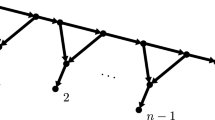
Content of \( treeset \) before the \(i^{\mathrm{th}}\) iteration of the loop in Algorithm 2 for \(n=10, n=11\) and \(n=12\). In case of \(n=11\), the tree with white leaves for \(i=7, \ldots , 10\), depicts the unique tree in \( treeset \) with an odd number of leaves. For \(n=10\), the leaf depicted as a diamond represents leaf u used in the proof of Lemma 10. Note that the tree containing this leaf is always clustered as late as possible. In case of \(n=12\), the leaf depicted as a diamond again represents leaf u used in the proof of Lemma 10. In this case, the tree containing this leaf is always clustered as soon as possible. The last tree depicted in each column represents the GFB tree. Note that \(T_n^{ gfb }\) can be obtained from \(T_{n-1}^{ gfb }\) by replacing the leaf depicted as a diamond by a cherry. Moreover, \(T_n^{ gfb }\) can be obtained from \(T_{n+1}^{ gfb }\) by replacing the cherry containing the diamond leaf by a single leaf
Now, first consider Algorithm 2 for \(n-1\), which is an even number. After the first \((n-3)/{2}\) iterations of the loop, \( treeset \) contains \((n-3)/{2}\) trees with 2 leaves and two trees with 1 leaf, which are clustered last to form the last cherry. We keep tracking one leaf u of this cherry throughout the algorithm. The algorithm at this stage contains only cherries, which are all isomorphic, so without loss of generality, we may assume that u is contained in the one that gets clustered with another tree last, i.e. after all other cherries have been clustered. We continue like this, always assuming without loss of generality (when there is more than one tree in \( treeset \) of the same size as the tree that contains u) that the tree containing u is in the last one to be clustered. By (i), this means that if we replace u in \(T_{n-1}^{ gfb }\) by a cherry, we derive \(T_{n}^{ gfb }\). This is due to the fact that in the analogous step where \( treeset \) for \(n-1\) only contains cherries, \( treeset \) for n will contain only cherries and a tree containing three leaves. This triplet will subsequently act like a cherry, but like the one that happens to be clustered last. So, we identify the cherry in this triplet with u to see the correspondence between \(T_{n-1}^{ gfb }\) and \(T_{n}^{ gfb }\). Note that this directly implies that \(T_{n-1}^{ gfb }\) and \(T_{n}^{ gfb }\) share a common maximal pending subtree—namely the one that does not contain u.
Note that by (ii), an analogous procedure for \(n+1\) leads to \(T_{n+1}^{ gfb }\) and \(T_{n}^{ gfb }\) sharing a common maximal pending subtree. In this case, we track a cherry in \(T_{n+1}^{ gfb }\), namely the one that happens to be clustered first, and replace it by a single leaf to see the correspondence between \(T_{n+1}^{ gfb }\) and \(T_{n}^{ gfb }\). This completes the proof. \(\square \)
We can proceed now to prove Proposition 5. Let \(n=2^m+p\) with \(m=\lfloor \log _2(n)\rfloor \) and \(0\leqslant p<2^m\). We shall prove by induction on n that if \(T_n^{ gfb }=(T_a,T_b)\) is a GFB tree with \(n\geqslant 2\), \(T_a\in {\mathcal {T}}_{n_a}\), \(T_b\in {\mathcal {T}}_{n_b}\) and \(n_a\geqslant n_b\) then:
- (i)
If \(0\leqslant p\leqslant 2^{m-1}\), \(n_a = 2^{m-1}+p\) and \(n_b = 2^{m-1}\) and then \(T_b\) is fully symmetric.
- (ii)
If \(2^{m-1}\leqslant p<2^m\), we have \(n_a = 2^{m}\) and \(n_b=p\) and then \(T_a\) is fully symmetric.
We want to point out that we understand that the conjunction of these two assertions in the case when both premises are satisfied, namely when \(p=2^{m-1}\), says that \(n_a =2^m\) and \(n_b = 2^{m-1}\) and then both \(T_a\) (by (ii)) and \(T_b\) (by (i)) are fully symmetric.
The base case for (i) is when \(n=2\) and for (ii), when \(n=3\). In both cases the assertions are obvious, because there is only one bifurcating tree with \(2=2^1+0\) leaves (a cherry with \(n_a=n_b=1=2^{0}\)) and only one bifurcating tree with \(3=2^1+1\) leaves (a caterpillar with \(n_a=2=2^1\) and \(n_b=1\)).
Now, let \(n \geqslant 4\) and assume that (i) and (ii) hold for up to \(n-1\) leaves. Let \(T=(T_a,T_b)\) be a GFB tree with n leaves, with \(T_a\in {\mathcal {T}}_{n_a}\), \(T_b\in {\mathcal {T}}_{n_b}\) and \(n_a\geqslant n_b\) Recall that \(T_a\) and \(T_b\) are again GFB trees by Lemma 5. We distinguish two cases, depending on the parity of n:
Assume that n is even, say \(n=2n_0\) with \(n_0\geqslant 2\). In this case, Algorithm 2 results in a tree \(T^{ gfb }_n\) with \(n_0\) cherries (because in each of the first \(n_0\) iterations of the loop a pair of nodes are merged into a cherry). We now consider the tree \(T'\) with \(n_0\) leaves that is obtained from \(T^{ gfb }_n\) by replacing all cherries by single leaves. Let \(T'=(T_a',T_b')\) be the decomposition into maximal pending subtrees, with \(T_a'\in {\mathcal {T}}_{n_a'}\), \(T_b'\in {\mathcal {T}}_{n_b'}\) and \(n_a'\geqslant n_b'\). By construction, \(T_a\) and \(T_b\) are obtained by replacing the leaves of \(T_a'\) and \(T_b'\) by cherries, and therefore, in particular, \(n_a=2n_a'\) and \(n_b=2n_b'\). Note now that, since \(T^{ gfb }_n\) is a GFB tree, so is \(T'\) (because as soon as Algorithm 2 only has cherries to choose from, they are treated like leaves). Note also that, since n is even, so is p, say \(p=2p_0\), and \(n_0=2^{m-1}+p_0\). Then we have that:
- (i)
If \(0\leqslant p\leqslant 2^{m-1}\), then \(0\leqslant p_0\leqslant 2^{m-1-1}\) and hence, by the induction hypothesis, \(n_a'=2^{m-2}+p_0\), \(n_b'=2^{m-2}\), and \(T_b'\) is fully symmetric, which implies that \(n_a=2n_a'=2^{m-1}+2p_0=2^{m-1}+p\), \(n_b=2n_b'=2^{m-1}\), and \(T_b\) is fully symmetric, because it is obtained from the fully symmetric tree \(T_b'\) by replacing all its leaves by cherries.
- (ii)
If \(2^{m-1}\leqslant p< 2^{m}\), then \(2^{m-1-1}\leqslant p_0\leqslant 2^{m-1}\) and hence, by the induction hypothesis, \(n_a'=2^{m-1}\), \(n_b'=p_0\), and \(T_a'\) is fully symmetric, which implies that \(n_a=2n_a'=2^{m}\), \(n_b=2n_b'=2p_0=p\), and, arguing as in (i), \(T_a\) is fully symmetric.
- (i)
Assume that n is odd, say \(n=2n_0+1\) with \(n_0\geqslant 2\). In this case both \(n-1=2n_0\) and \(n+1=2(n_0+1)\) are even. Write \(n=2^m+p\) and \(p=2p_0+1\), so that \(n_0=2^{m-1}+p_0\) with \(0\leqslant p_0<2^{m-1}\). Let \(T^1 :=T_{n-1}^{ gfb }\) and \(T^2:=T_{n+1}^{ gfb }\). The tree \(T^1\) satisfies (i) and (ii) by the induction hypothesis, and it can be proved that \(T^2\) also satisfies these assertions by arguing as in the previous case when n is even (i.e. replacing the pending \(n_0+1\) cherries in \(T^2\) by single leaves, noticing that the resulting tree is GFB, applying the induction hypothesis to it and finally returning back to \(T^2\) by replacing leaves by cherries). Let \(T^1=(T^1_a, T^1_b)\)—with \(T^1_a\in {\mathcal {T}}_{n^1_a}\) and \(T^1_b\in {\mathcal {T}}_{n^1_b}\) and \(n^1_a\geqslant n^1_b\)—and \(T^2=(T^2_a, T^2_b)\)—with \(T^2_a\in {\mathcal {T}}_{n^2_a}\) and \(T^2_b\in {\mathcal {T}}_{n^2_b}\) and \(n^2_a\geqslant n^2_b\)—denote the decompositions of \(T^1\) and \(T^2\) into maximal pending subtrees, respectively. Note that, since n is odd, \(p\ne 0,2^{m-1}\). Now we have:
- (i)
If \(0< p<2^{m-1}\), then \(n-1=2^m+(p-1)\) with \(0\leqslant p-1<2^{m-1}\) and \(n+1=2^m+(p+1)\) with \(0<p+1\leqslant 2^{m-1}\). Then, since \(T^1\) and \(T^2\) satisfy assertion (i),
$$\begin{aligned} n^1_a=2^{m-1}+p-1,\ n^1_b=2^{m-1},\ n^2_a=2^{m-1}+p+1,\ n^2_b=2^{m-1} \end{aligned}$$and both \(T_b^1\) and \(T_b^2\) are fully symmetric and hence (since they have the same numbers of leaves) \(T_b^1=T_b^2\). Now, we know by Lemma 10 that T shares a maximal pending subtree with \(T^1\) and a maximal pending subtree with \(T^2\). Looking at the numbers of leaves of the maximal pending subtrees of \(T^1\) and \(T^2\), one easily deduces that the only possibility for this to happen is that T shares with \(T^1\) and \(T^2\) the same maximal pending subtree: the fully symmetric subtree \(T_b^1=T_b^2\). (Indeed, since \(T_a^1\ne T_a^2\), because they have different numbers of leaves, if T did not share \(T_b^1=T_b^2\) with both \(T^1\) and \(T^2\), then it would have a maximal pending subtree in common with \(T^1\) and the other maximal pending subtree in common with \(T^2\), but no combination of a maximal pending subtree of \(T^1\) and a maximal pending subtree of \(T^2\) yields a tree with \(2^m+p\) leaves.) A fortiori, one of the maximal pending subtrees of T is a fully symmetric tree with \(2^{m-1}\) leaves and the other must have, thus, the remaining \(2^{m-1}+p\) leaves. This shows that \(n_a=2^{m-1}+p\) and \(n_b=2^{m-1}\) and \(T_b\) is fully symmetric.
- (ii)
If \(2^{m-1}<p\leqslant 2^{m}-3\) then \(n-1=2^m+(p-1)\) with \(2^{m-1}\leqslant p-1<2^{m}\) and \(n+1=2^m+(p+1)\) with \(2^{m-1}<p+1< 2^{m}\). Then, since \(T^1\) and \(T^2\) satisfy assertion (ii),
$$\begin{aligned} n^1_a=2^{m},\ n^1_b=p-1,\ n^2_a=2^{m},\ n^2_b=p+1 \end{aligned}$$and both \(T_a^1\) and \(T_a^2\) are fully symmetric and hence (since they have the same numbers of leaves) \(T_a^1=T_a^2\). Reasoning as in the previous case, we deduce that T shares with both \(T^1\) and \(T^2\) the fully symmetric maximal pending subtree \(T_a^1=T_a^2\). In particular, one of its maximal pending subtrees has \(2^{m}\) leaves (and it is fully symmetric) and the other must have, thus, the remaining p leaves. This shows that \(n_a=2^{m}\) and \(n_b=p\) and \(T_a\) is fully symmetric.
- (iii)
Consider finally the case when \(p= 2^{m}-1> 2^{m-1}\). Then, \(n-1=2^m+(p-1)\) with \(2^{m-1}\leqslant p-1<2^{m}\) and \(n+1=2^{m+1}\). In this case, since \(T^1\) satisfies assertion (ii) and \(T^2\) satisfies assertion (i),
$$\begin{aligned} n^1_a=2^{m},\ n^1_b=2^{m}-2,\ n^2_a=2^{m},\ n^2_b=2^{m} \end{aligned}$$and \(T_a^1\), \(T_a^2\) and \(T_b^2\) are fully symmetric and hence (since they have the same numbers of leaves) \(T_a^1=T_a^2=T_b^2\). Arguing as in the previous cases we conclude that T has a maximal pending subtree with \(2^m\) leaves that is fully symmetric and the other maximal pending subtree with the remaining \(2^m-1\) leaves, and hence it satisfies assertion (ii).
- (i)
This completes the proof.
1.3 A.3 Proof of Proposition 8
This appendix is devoted to establish the following result.
Proposition 8
For every \(n\geqslant 1\), let \(n=\sum _{i=1}^\ell 2^{m_i}\), with \(\ell \geqslant 1\) and \(m_1>\cdots > m_\ell \), be its binary expansion.
- (a)
\(s(T_n^ gfb )=n-1-(m_1-m_\ell )\).
- (b)
For every \(T\in \widetilde{\mathcal {MC}}_n\), if \(T\ne T_n^ gfb \), then \(s(T)< s(T_n^ gfb )\).
Proof
Note first of all that the number s of symmetry vertices satisfies the following recurrence: if \(T\in {\mathcal {T}}_1\), then \(s(T)=0\), and if \(T=(T_a,T_b)\in {\mathcal {T}}_n\) with \(n\geqslant 2\), then
We shall now prove (a) by induction on n. When \(n=1=2^0\), the statement holds because \(s(T_1^ gfb )=0 = 1-1-(0-0) = n-1-(m_1-m_\ell )\). More in general, the statement clearly holds whenever n is a power of 2, say \(n=2^{m_1}\), because in this case \(T^{ gfb }_n\) is fully symmetric and therefore all its internal nodes are symmetry vertices, i.e. \(s(T^{ gfb }_n)=n-1=n-1-(m_1-m_1)\).
Now assume that the statement holds for every GFB tree with \(n'\) leaves, with \(n'<n\), and consider the tree \(T^{ gfb }_n\). By Lemma 5, if \(T^{ gfb }_n= (T_a, T_b)\), then \(T_a\) and \(T_b\) are GFB trees and, by the inductive hypothesis, the statement holds for \(T_a=T_{n_a^ gfb }^ gfb \) and \(T_b=T_{n_b^ gfb }^ gfb \).
Let us now write n as \(2^m+p\) with \(m = \lfloor \log _2(n) \rfloor \) and \(0 \leqslant p < 2^m\), and consider its binary expansion \(n = \sum _{j=1}^\ell 2^{m_j}\) with \(m_1>\cdots >m_\ell \), so that \(m_1=m\) and \(p = \sum _{j=2}^\ell 2^{m_j}\) is the binary expansion of p if \(p>0\). Now, we distinguish four cases:
- (i)
If \(p=0\), then n is a power of 2, in which case we have already seen that the statement holds.
- (ii)
If \(1\leqslant p<2^{m-1}\), then, by Proposition 5, \(n_a^ gfb = 2^{m-1} + p\) and \(n_b^ gfb = 2^{m-1}\) and \(T_b\) is fully symmetric. In this case, \(m_2<m-1=m_1-1\) and thus \(n_a^ gfb = 2^{m_1 - 1} + \sum _{j=2}^\ell 2^{m_j}\) is the binary expansion of \(n_a^ gfb \). Then \(s(T_b)=2^{m-1}-1\) and, by the induction hypothesis,
$$\begin{aligned} s(T_a)=2^{m-1} + p-1-(m_1-1-m_\ell )=2^{m-1} + p-(m_1-m_\ell ) \end{aligned}$$and hence
$$\begin{aligned} s(T^{ gfb }_n)= & {} s(T_a) + s(T_b)= 2^{m-1} + p-(m_1-m_\ell ) + 2^{m-1}-1\\= & {} n- 1- (m_1 - m_\ell ). \end{aligned}$$ - (iii)
If \(p= 2^{m-1}\), so that \(n=2^m+2^{m-1}\) is the binary expansion of n, then, by Proposition 5, \(n_a^ gfb = 2^{m}\) and \(n_b^ gfb =2^{m-1}\) and both \(T_a\) and \(T_b\) are fully symmetric. In this case, \(s(T_a)=2^m-1\) and \(s(T_b)=2^{m-1}-1\) and hence
$$\begin{aligned} s(T^{ gfb }_n)&= s(T_a) + s(T_b)= 2^m-1 + 2^{m-1}-1 \\&= 2^m + 2^{m-1} - 1 - (m-(m-1)) = n-1-(m_1-m_\ell ). \end{aligned}$$ - (iv)
Finally, assume that \(p > 2^{m-1}\), so that its binary expansion is \(p= 2^{m-1} + \sum _{i=3}^\ell 2^{m_i}\), and in particular \(m_2=m-1=m_1-1\). In this case, by Proposition 5, \(n_a^ gfb = 2^m\), and \(T_a\) is fully symmetric, and \(n_b^ gfb = p\). Then, \(s(T_a)=2^m-1\) and, by the induction hypothesis, \(s(T_b)=p-1-(m_1-1-m_\ell )=p-(m_1-m_\ell )\) and hence
$$\begin{aligned} s(T) = s(T_a) + s(T_b) = 2^{m}-1+p-(m_1-m_\ell )= n-1-(m_1 - m_\ell ). \end{aligned}$$
This completes the proof of (a).
As far as (b) goes, we also prove it by induction on n. The case \(n=1\) is obvious, since there is only one bifurcating tree in \({\mathcal {T}}_1\). Let now \(n\geqslant 2\) and assume that the statement is true for every number \(n'\) of leaves smaller than n. Let \(T = (T_a,T_b)\), with \(T_a\in {\mathcal {T}}_{n_a}\), \(T_b\in {\mathcal {T}}_{n_b}\), and \(n_a\geqslant n_b\), be a minimal Colless tree with n leaves such that s(T) is maximum in \(\widetilde{\mathcal {MC}}_n\). We want to prove that \(T=T^{ gfb }_n\).
By Lemma 2, \(T_a\in \widetilde{\mathcal {MC}}_{n_a}\) and \(T_b\in \widetilde{\mathcal {MC}}_{n_b}\) and therefore, by the inductive hypothesis, \(s(T_a)\leqslant s(T_{n_a}^ gfb )\) and \(s(T_b)\leqslant s(T_{n_b}^ gfb )\). To prove that \(T=T^{ gfb }_n=(T_{n_a^ gfb }^ gfb ,T_{n_b^ gfb }^ gfb )\), it is enough to prove that \(n_a=n_a^ gfb \) and \(n_b=n_b^ gfb \) (and, actually, it is enough to prove one of these equalities, because then the other will follow from \(n_a+n_b=n=n_a^ gfb +n_b^ gfb \)) and that \(s(T_a)= s(T_{n_a}^ gfb )\) and \(s(T_b)= s(T_{n_b}^ gfb )\) (because by the inductive hypothesis these equalities imply that \(T_a=T_{n_a}^ gfb \) and \(T_b=T_{n_b}^ gfb \)). Let \(n_a=\sum _{i=1}^{\ell _a} 2^{s_i}\) and \(n_b=\sum _{i=1}^{\ell _b} 2^{t_i}\) be the binary decompositions of \(n_a\) and \(n_b\).
Now, two cases arise, depending on whether the root of T is a symmetry vertex or not. Let us assume first that it is a symmetry vertex, i.e, that \(T_a=T_b\). In this case, n must be even and \(n_a = n_b = n/2=\sum _{i=1}^{\ell } 2^{m_i-1}\). In particular \(s_1=t_1=m_1-1\) and \(s_{\ell _a}=t_{\ell _b}=m_{\ell }-1\). Moreover, it must happen that \(s(T_a)= s(T_{n/2}^ gfb )\), because if \(s(T_a)< s(T_{n/2}^ gfb )\) and if we denote by \(T'\) the tree \((T_{n/2}^ gfb ,T_{n/2}^ gfb )\), then \(T'\in \widetilde{\mathcal {MC}}_n\) by Proposition 1 (recall that (n/2, n/2) always belongs to QB(n)) and, by Eq. (19),
against the assumption that s(T) is maximum in \(\widetilde{\mathcal {MC}}_n\). So, in this case it remains to prove that \(n_a^ gfb =n_b^ gfb =n/2\).
Now, applying Eq. (19) and (a), we have that
Thus, if \(\ell >1\), then \(s(T)<s(T_n^ gfb )\), against the assumption that s(T) is maximum in \(\widetilde{\mathcal {MC}}_n\). Therefore, \(\ell =1\), i.e. \(n=2^{m_1}\) and hence \(n_a^ gfb =n_b^ gfb =n/2=n_a=n_b\), as we wanted to prove.
Let us assume now that the root of T is not a symmetry vertex. Recall from Corollary 9 that
Combining these inequalities with Proposition 5 we obtain that:
If \(0\leqslant p< 2^{m_1-1}\), then
$$\begin{aligned} 2^{m_1-1}=n_b^ gfb \leqslant n_b\leqslant n_a\leqslant n_a^ gfb = 2^{m_1-1}+p<2^{m_1}, \end{aligned}$$(20)and then, in this case, \(s_1=t_1=m_1-1\).
If \(2^{m_1-1}\leqslant p<2^{m_1}\), then
$$\begin{aligned} 2^{m_1-1}\leqslant p =n_b^ gfb \leqslant n_b\leqslant n_a\leqslant n_a^ gfb =2^{m_1} \end{aligned}$$(21)and then either \(n_a=2^{m_1}=n_a^ gfb \), in which case \(n_b=p=n_b^ gfb <2^{m_1}\), \(s_1=m_1\), and \(t_1=m_1-1\), or \(2^{m_1-1}\leqslant n_b\leqslant n_a<n_a^ gfb =2^{m_1}\), in which case \(s_1=t_1=m_1-1\).
So, in particular, \(t_1\) is always \(m_1-1\), and \(s_1\) is \(m_1\), when \(n_a=2^{m_1}=n_a^ gfb \), and \(m_1-1\) otherwise. Moreover, since \(n_a+n_b=n\), it always happens that \(\min \{s_{\ell _a}, t_{\ell _b}\} \leqslant m_\ell \).
Now, in this case we have again that \(s(T_a)=s(T_{n_a}^ gfb )\) and \(s(T_b)=s(T_{n_b}^ gfb )\), because if, say, \(s(T_a)<s(T_{n_a}^ gfb )\) and if we replace in T its maximal pending subtree \(T_a\) by \(T_{n_a}^ gfb \), then, by Proposition 1 (and recalling that, since \(T\in \widetilde{\mathcal {MC}}_n\), by that very proposition we have that \((n_a,n_b)\in QB(n)\)), the resulting tree \(T'=(T_{n_a}^ gfb ,T_b)\) is still minimal Colless and, by Eq. (19),
against the assumption that s(T) is maximum in \(\widetilde{\mathcal {MC}}_n\). So, \(T_a=T_{n_a}^ gfb \) and \(T_b=T_{n_b}^ gfb \). It remains to prove that \(n_a=n_a^ gfb \) and \(n_b=n_b^ gfb \).
By Eq. (19) and (a), we have that
We consider now several possibilities:
If \(s_{\ell _a}=s_1\), then \(n_a=2^{s_1}\), where \(s_1\) is \(m_1-1\) or \(m_1\). Now, since \(2^{m_1-1}\leqslant n_b\leqslant n_a\) and \(n_a+n_b=2^{m_1}+p\), if we had \(n_a=2^{m_1-1}\), we would also have \(n_b=2^{m_1-1}\) and \(p=0\), and then \(n_a^ gfb =n_a=n_b=n_b^ gfb \); but then \(T_a=T_{n_a}^ gfb \) and \(T_b=T_{n_b}^ gfb \) would be isomorphic to the fully symmetric tree \(T_{m_1-1}^ fs \) and hence the root of T would be a symmetry vertex, against the current assumption that it is not so.
So, in this case we have \(n_a=2^{m_1}\). By properties (20) and (21), it can only happen when \(2^{m_1-1}\leqslant p\) and \(n_a= n_a^ gfb \), and then \(n_b= n_b^ gfb \), too.
If \(s_{\ell _a}<s_1\) and \(t_{\ell _b}\leqslant s_{\ell _a}\), then \(t_{\ell _b}=\min \{s_{\ell _a}, t_{\ell _b}\} \leqslant m_\ell \) and hence, by (22),
$$\begin{aligned} s(T)=s(T_{n}^ gfb )-(s_1+m_\ell -s_{\ell _a}-t_{\ell _b})<s(T_{n}^ gfb ), \end{aligned}$$against the assumption that s(T) is maximum among all minimal Colless trees with n leaves.
If \(s_{\ell _a}<s_1\) and \(s_{\ell _a}<t_{\ell _b}\), then \(s_{\ell _a}=\min \{s_{\ell _a}, t_{\ell _b}\} \leqslant m_\ell \). Since in this case \(n_a\) is not a power of 2, we have \(s_1=t_1=m_1-1\) and then
$$\begin{aligned} s_1+m_\ell -s_{\ell _a}-t_{\ell _b}\geqslant m_1-1-t_{\ell _b}=t_1-t_{\ell _b}\geqslant 0. \end{aligned}$$If one of these inequalities is strict, we deduce again that
$$\begin{aligned} s(T)=s(T_{n}^ gfb )-(s_1+m_\ell -s_{\ell _a}-t_{\ell _b})<s(T_{n}^ gfb ), \end{aligned}$$reaching the same contradiction as before. Therefore, both inequalities are equalities and hence \(s_{\ell _a}=m_\ell \) and \(t_{\ell _b}=t_1=m_1-1\), which implies in particular that \(n_b=2^{m_1-1}\). By (20) and (21), this can only happen when \(0\leqslant p\leqslant 2^{m_1-1}\) and \(n_b=2^{m_1-1}=n_b^ gfb \).
This finishes the proof of (b). \(\square \)
1.4 A.4 Computation of some probabilities under the \(\beta \)-model
Aldous’ \(\beta \) model (Aldous 1996) is a probabilistic model of bifurcating phylogenetic trees that depends on one parameter \(\beta \in (-2,\infty )\). As any other such probabilistic model, it yields a probabilistic model of bifurcating unlabeled trees, by defining the probability \(P_\beta (T)\) of a tree \(T\in {\mathcal {T}}_n\) as the sum of the probabilities of all phylogenetic trees on n leaves with shape T. This probabilistic model of bifurcating unlabeled trees satisfies the following Markovian recurrence. For every \(m\geqslant 2\) and \(k=1,\ldots ,m-1\), let
where \(a_m(\beta )\) is a suitable normalizing constant so that \(\sum \limits _{a=1}^{m-1} q_{m,\beta }(a)=1\), and
Then, if \(T=(T_{a},T_b)\in {\mathcal {T}}_n\) with \(T_a\in {\mathcal {T}}_{n_a}\), \(T_b\in {\mathcal {T}}_{n_b}\) and \(n_a\geqslant n_b\),
Recall that the Gamma function \(\varGamma \) satisfies the recurrence \(\varGamma (x+1)=x\varGamma (x)\) and that, for every \(n\in {\mathbb {N}}_{\geqslant 1}\), \(\varGamma (n)=(n-1)!\).
We want to compute the probabilities under this model of \(T_6^ mb \) and \(T_6^ gfb \). To do that, we shall need to compute all values \(q_{6,\beta }(k)\) (we need all of them in order to compute the normalizing constant \(a_6(\beta )\)):
Imposing now \(\sum _{k=1}^5 q_{6,\beta }(k)=1\), i.e.
and solving for \(a_6(\beta )\), we obtain
We can compute now the desired probabilities:
As far as \(P_\beta (T_6^ mb )\) goes, by Eq. (23) we have that
$$\begin{aligned} P_\beta (T_6^ mb )=q_{6,\beta }(3)\cdot P_\beta (T_3^ mb )^2=q_{6,\beta }(3) \end{aligned}$$because \({\mathcal {T}}_3=\{T_3^ mb \}\) and hence \(P_\beta (T_3^ mb )=1\). So,
$$\begin{aligned} P_\beta (T_6^ mb )&= \frac{3\cdot 5!\cdot (\beta +3)^2(\beta +2)^2\varGamma (\beta +2)^2}{3!^2(\beta +3)(\beta +2)\varGamma (\beta +2)^2(31 \beta ^2 + 194\beta + 300)}\\&= \frac{10 (\beta +3)(\beta +2)}{31 \beta ^2 + 194\beta + 300}. \end{aligned}$$As to \(P_\beta (T_6^ gfb )\) goes, by Eq. (23) (and the fact that \(T_6^ gfb = (T_4^ gfb , T_2^ gfb )\) by Lemma 5 and Proposition 5) we have that
$$\begin{aligned} P_\beta (T_6^ gfb )=2q_{6,\beta }(4)\cdot P_\beta (T_2^ gfb )P_\beta (T_4^ gfb ) \end{aligned}$$where \(P_\beta (T_2^ gfb )=1\), because \(T_2^ gfb \) is the only tree in \({\mathcal {T}}_2\);
$$\begin{aligned} P_\beta (T_4^ gfb )= P_\beta (T_4^ mb )=\frac{3(\beta +2)}{7\beta +18} \end{aligned}$$by Lemma 4 in (Coronado et al. 2019); and
$$\begin{aligned} q_{6,\beta }(4)&= \frac{3\cdot 5!\cdot (\beta +4)(\beta +3)(\beta +2)^2\varGamma (\beta +2)^2}{2\cdot 4!\cdot (\beta +3)(\beta +2)\varGamma (\beta +2)^2(31 \beta ^2 + 194\beta + 300)}\\&= \frac{15 (\beta +4)(\beta +2)}{2(31 \beta ^2 + 194\beta + 300)}. \end{aligned}$$So, finally,
$$\begin{aligned} P_\beta (T_6^ gfb )&=2\cdot \frac{15 (\beta +4)(\beta +2)}{2(31 \beta ^2 + 194\beta + 300)}\cdot \frac{3(\beta +2)}{7\beta +18}\\&= \frac{45 (\beta +4)(\beta +2)^2}{(31 \beta ^2 + 194\beta + 300)(7\beta +18)}. \end{aligned}$$
Rights and permissions
About this article

Cite this article
Coronado, T.M., Fischer, M., Herbst, L. et al. On the minimum value of the Colless index and the bifurcating trees that achieve it. J. Math. Biol. 80, 1993–2054 (2020). https://doi.org/10.1007/s00285-020-01488-9
Received:
Revised:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00285-020-01488-9