
Abstract
Studies of cyanobacterial nostocacean taxa are important to the global scientific community, mainly because a significant number of beneficial strains that belong to the order Nostocales fix atmospheric nitrogen, thus contributing to the fertility of agricultural soils worldwide, while others behave as nuisance microorganisms in aquatic ecosystems due to their involvement in toxic bloom events. However, in spite of their ecological importance and environmental concerns, their identification and taxonomy are still problematic and doubtful, often being based on current morphological and physiological studies, which generate confusing classification systems and usually vary under different conditions. Therefore, the present research aimed to investigate through a polyphasic approach differences in morphological, biochemical and genotypic features of three nostocacean cyanobacterial strains isolated from central-western Portuguese shallow freshwater bodies. Morphometric, genetic (16S rRNA, nifH and hetR fragments) and biochemical (fatty acid methyl ester; FAME profiles) data were used to characterize the strains. Morphological analysis and sequencing of 16S rRNA fragments showed that the strains belonged to Anabaena cylindrica (UTAD_A212), Aphanizomenon gracile (UADFA16) and Nostoc muscorum (UTAD_N213) species. These strains showed clear distinct morphological and genetic features, allowing easy allocation to their respective genera. The same happened by using partial sequences of hetR and nifH genes, in spite of the scarcity of deposited sequences. Biochemical characterization showed that the FAME profiles obtained were consistent with both morphological and molecular analyses. It was suggested that the ratio of monounsaturated to polyunsaturated FAMEs, together with the unsaturation index, could be used as genus-specific chemotaxonomic biomarkers.
Similar content being viewed by others
Introduction
Cyanobacteria are impressive ecosystem engineers with an evolutionary history stretching back at least 2.15 billion years (Hayes et al., 2007; Rasmussen et al., 2008). These oxygenic photoautotrophic prokaryotes are widely distributed in natural environments and constitute a major component of microbial populations in terrestrial and aquatic habitats worldwide. They are often referred to as ‘miniature factories’ of the biological world and represent an alternative source of a variety of bioactive compounds, lipids/fatty acids, proteins, enzymes, pigments and compounds of pharmaceutical and nutraceutical value (Schaeffer & Krylov, 2000; Rastogi & Sinha, 2009).
According to Castenholz (1989) and Komárek & Anagnostidis (1989), the order Nostocales includes filamentous cyanobacteria that are capable of cell differentiation in heterocysts, akinetes or reproductive trichomes (hormogonia). The genera Anabaena, Aphanizomenon and Nostoc belong to the family Nostocaceae by traditional classification (Komárek & Anagnostidis, 1989; Komárek, 2010) and subsection IV according to bacteriological classification (Rippka et al., 1979; Castenholz, 1989). This family is characterized morphologically by (i) isopolar filaments, (ii) absence of any branching (with the exception of certain anomalies), (iii) presence of heterocysts (with the exception of secondary derived genotypes) and (iv) facultative presence of typical paraheterocytic or apoheterocytic akinetes (Komárek, 2010).
Information on cyanobacterial taxa present in rice fields and other agro-ecosystems in Europe is still scarce and differs significantly from that obtained in Asiatic countries, especially concerning different climatic conditions and pesticide application methodologies (Ariosa et al., 2006; Zancan et al., 2006). The native strains frequently found in European rice fields mainly belong to the heterocystous genera Anabena and Nostoc, but also Calothrix and Tolypothrix (Whitton, 2000; Ariosa et al., 2006; Galhano et al., 2009). On the other hand, the diversity, ecology and genetic heterogeneity among planktonic Anabaena-like and Aphanizomenon-like morphospecies require a deeper characterization, since there is neither a clear phylogenetic nor a morphological separation (Hindák, 2000; Rajaniemi et al., 2005; Stüken et al., 2009). The few existing studies on taxonomy involving planktonic Aphanizomenon and Anabaena strains from Southern Europe were carried out by Pereira et al. (2004), Quesada et al. (2006) and de Figueiredo et al. (2010), while for Northern Europe the number of publications is vast (Hindák, 2000; Lyra et al., 2001; Gugger et al., 2002a; Rajaniemi et al., 2005; Zwart et al., 2005; Komárek & Komárková, 2006; Willame et al., 2006; Rücker et al., 2007; Stüken et al., 2009; Zapomělová et al., 2010).
The taxonomy of cyanobacteria has been traditionally based on morphological, physiological and ecological characteristics, but recently also on biochemical and molecular approaches (Gugger et al., 2002b; Rajaniemi et al., 2005; Hayes et al., 2007; Nayak et al., 2007; Li et al., 2008; Zapomělová et al., 2010). Molecular techniques based on PCR amplification targeting conserved regions inside the 16S rRNA gene have allowed determination of phylogenetic affiliations among cyanobacteria and development of modern cyanobacterial taxonomy (Komárek, 2006). Other genes coding for proteins, such as hetR (Han et al., 2009; Schleifer, 2009) and nifH (Zehr & McReynolds, 1989; Janson et al., 1998), have also been shown to have phylogenetic discriminating potential. However, many discrepancies between traditional morphological classification (Rippka et al., 1979; Castenholz, 1989; Castenholz & Waterbury, 1989) and phylogenetic analysis are still unresolved (Gugger et al., 2002a, b; Komárek, 2010). Therefore, chemotaxonomic markers such as lipids and their fatty acids have been considered as complementary methodologies, since they provide interesting information for taxonomic position assignment and some correlations with morphological properties of cyanobacteria (Wilmotte, 1994; Li & Watanabe, 2001, 2004; Gugger et al., 2002a, b; Temina et al., 2007; Li et al., 2008). Notwithstanding, the literature on lipid composition of freshwater cyanobacteria inhabiting different natural ecosystems in Portugal is non-existent and polyphasic studies are scarce.
In the present study, molecular information regarding the 16S rRNA gene as well as the hetR and nifH genes was obtained from filamentous cyanobacterial strains isolated from two Portuguese freshwater ecosystems, namely rice fields (genera Anabaena and Nostoc) and a eutrophic shallow lake (genus Aphanizomenon). The genotypic features, together with morphological and biochemical attributes (fatty acid methyl ester profiles), were used to characterize the strains.
Materials and methods
Cyanobacterial isolation and culture conditions
The strains Anabaena cylindrica UTAD_A212 and Nostoc muscorum UTAD_N213 were originally collected in the summer of 2005 (mid-August) from pesticide-free rice field soils located near Montemor-o-Velho in the Mondego River basin (40°10′N, 8°41′W, central-western Portugal). The choice of these strains was based on their higher abundance relatively to other local native rice field cyanobacteria, thus making them promising N2-fixing microorganisms in future rice biofertilization programmes. The strain Aphanizomenon gracile UADFA16 was isolated from the eutrophic shallow Vela Lake near Quiaios village, Figueira-da-Foz (44°58′N, 5°18′W, central-western Portugal) in the summer of 2007, and was chosen to infer and clarify taxonomic relationships with other studied strains. The study area details of sampled rice fields were similar to those described by Marques et al. (2008), and Vela Lake biotic and abiotic characteristics were extensively monitored previously (de Figueiredo et al., 2006). The samples from rice fields and Vela Lake were collected in situ according to the methodologies described in Galhano et al. (2009) and de Figueiredo et al. (2006), respectively. Cyanobacterial strains were isolated by capillarity, agar plate spreading and serial dilution techniques (Andersen & Kawachi, 2005). Following the preparation of axenic and unialgal cultures by standard microbiological techniques (Guillard, 2005), strains were cultivated in liquid BG110 medium (pH 7.4) without sodium nitrate (Rippka et al., 1979), with minor modifications. Axenic maintenance of the cultures was periodically checked by careful optical microscopy examination, and sometimes cycloheximide (Actidione) at 70 mg l−1 was added to inhibit eukaryotic cell growth. Strains grew for a period of 3–4 weeks in a rotary shaker (100 rpm) inside a culture growth chamber (Conviron mod. E7/2) at 28 ± 2°C and illuminated under a 16:8 h light–dark cycle, with an average photon flux density of 50 μmol m−2 s−1. Cultures were also shaken by hand two to four times daily to prevent cell clumping, mainly in N. muscorum UTAD_N213 flasks. Uniform suspensions of axenic clonal cultures for the experiments were maintained in exponential growth phase (14–21 days) by periodically subculturing them, and were grown in 500 ml cotton-plug closed Erlenmeyer flasks containing 250 ml of freshly autoclaved BG110. Growth was measured as OD750 in an UV/VIS Varian mod. Cary 100 spectrophotometer.
Morphological characterization and identification
For morphological analysis, cell dimensions (length and width of the vegetative cells, heterocysts and akinetes) of different filaments of each strain from late exponential growing fresh cultures were measured. Regular visual examination was carried out in N-free liquid and solid (agar) BG110 media. The presence of akinetes was scarce after the isolation of strains from their natural habitats, and was detected with accuracy only in old laboratory cultures, and sometimes only after a severe stress exposure, e.g., against xenobiotics (Galhano et al., 2009). Examination was carried out in an inverted research microscope (Olympus mod. IX71; Olympus Corporation, Tokyo) with a coupled digital camera (CCD Camera, ColorView III; Soft Imaging System GmbH, Münster), and CELL* ver. 3.2 image analysis software (Soft Imaging System GmbH, Münster). Photomicrographs were taken before morphometric determination in the exponential phase of growth. Taxonomic identification was based on morphological characteristics presented in the ‘classical’ standard literature (Geitler, 1932; Desikachary, 1959; Rippka et al., 1979; Castenholz, 1989; Komárek & Anagnostidis, 1989) and in more recent taxonomic revisions (Hindák, 2000; Rajaniemi et al., 2005; Komárek & Komárková, 2006; Willame et al., 2006; Komárek, 2010).
Extraction of total fatty acids
Cells were harvested by centrifugation (speed and time adjusted to each strain) at late exponential phase for fatty acid analysis, concentrated 10 times in 1-ml Eppendorf tubes and stored at −80°C until required. Cell samples were thawed, washed three times in BG110 medium, filtered and briefly ultrasonicated (30 s, using a Vibra-cell sonicator mod. VCX 130, Sonics Materials, equipped with a 3 mm diameter probe operating at 80%, in 5 s pulses) immediately before lipid extraction. The extraction of total lipids from cyanobacterial cells was done according to Sato & Murata (1988) with modifications, using successive vortex-mixing and vigorous shaking in teflon-lined screw-capped glass test tubes followed by centrifugation. Extractions were repeated until negligible amounts of pigment were extracted. The CHCl3 phase was vortex-mixed with a double volume of KCl solution (0.1 M). After centrifuging for 5 min at 3,000 rpm, the lower phase was collected using a glass Pasteur pipette and dried under a stream of N2. For the preparation of fatty acid methyl esters (FAMEs), a solution of 5% H2SO4 in CH3OH was used as the derivatizing reagent at 70°C for 2 h. The test tube contents were cooled, diluted with water and extracted with 5 ml of n-hexane (p.a. chromatographic grade), containing 0.01% of butylated hydroxytoluene (BHT, Sigma–Aldrich, St. Louis, USA) as antioxidant agent. The different lipid classes were transmethylated after addition of heptadecanoic acid (C:17, 99%, CAS No. 506-12-7, Fluka/Sigma-Aldrich, Steinheim, Germany) as internal standard. The n-hexane layer was dried over anhydrous Na2SO4 and evaporated to dryness. The final FAME residue was then dissolved in 400 μl n-hexane, kept in a nitrogen atmosphere and stored at −20°C under airtight GC-vials until analysis.
Gas chromatographic analysis of fatty acids
Whole-cell FAMEs were analysed with a gas chromatograph (Trace GC 2000, Thermo-Finnigan; Thermo Electron Corporation, Austin TX, USA) with helium as the carrier gas (flow rate of 40 cm s−1 at 140°C). A 4 μl sample was injected in split injection mode. Solvent blanks were checked periodically for impurities. We used an Agilent J&W Scientific (Agilent Technologies, Inc., USA) capillary column (DB-225MS; 30 m × 0.25 mm ID × 0.25 μm film thickness); the temperature programme of the GC oven was 2 min at 140°C, with a casting ramp of 4°C/min until 220°C, this temperature being maintained for 5 min. The temperature of the injector and FID was kept at 250 and 270°C, respectively. FAMEs were identified by co-chromatography of known authentic standards (37-Component FAME mix, ref. G003461 from Sigma-Aldrich, USA). The relative concentrations of different FAMEs were calculated by the percentage area method without correction for response factor, comparing the peak areas of their corresponding FAMEs with that of the internal standards cited above. Quantification was also accomplished by proper normalization against C:17. Peak area integration was done by using appropriate data processor software (Qual Browser/Xcalibur ver. 1.4 SR1, Thermo Electron Corp., San Jose, CA 95134, USA). Only those peaks listed in Fig. 2 could be identified beyond doubt. A fatty acid shorthand notation in the form of A:BωX was used, where A represents the number of carbon atoms, B is the number of double bonds and X gives the position of the double bond closest to the terminal methyl group. The unsaturation index (UI) was calculated according to the formula: UI = Σ % of unsaturated fatty acids × number of double bonds of each unsaturated fatty acid (Rosa & Catalá, 1998).
DNA extraction and PCR amplifications
The cyanobacterial DNA was extracted after centrifugation of 2 ml of the culture, resuspension in 200 μl of TE buffer (10 mM Tris–HCl, 1 mM ethylenediamine tetraacetic acid (EDTA), pH 8.0), lysis by adding 1 mg ml−1 lysozyme and incubation at 37°C for 1 h. Genomic DNA Purification Kit (MBI Fermentas, Vilnius, Lithuania) was used for DNA extraction and purification. DNA was resuspended in TE buffer and stored at −20°C. PCR amplification of 16S rRNA gene fragments was performed by combining the bacterial universal primer 27F (Lane, 1991) with the cyanobacteria-specific reverse primer CYA781R (Nübel et al., 1997), and the cyanobacteria-specific forward primers CYA106F (Nübel et al., 1997) and CYA738F (Valério et al., 2005) along with the bacterial universal primer 1492R (Lane, 1991). A fragment of the nifH gene (approximately 330 bp) was amplified by using degenerate primers (Zehr & McReynolds, 1989) and the hetR gene fragment (of about 450 bp) was obtained by using the primers hetr1 and hetr2 (Janson et al., 1998). Primers were commercially synthesized (STABVida, Oeiras, Portugal). PCRs were performed in a Bio-Rad iCycler Thermal Cycler (Hercules, California, USA) with 50 μl reaction mixtures each containing 3 mM MgCl2, 200 μM of each nucleotide, 1× PCR buffer with (NH4)2SO4, 5% dimethylsulfoxide (DMSO), 15 pmol of each primer, 1 U of Taq DNA polymerase and 50–200 ng template DNA. The PCR programmes for the primer set targeting the 16S rRNA gene fragment had an initial denaturation step at 94°C for 5 min, followed by 30 cycles consisting of 1 min at 94°C, 1 min at 50°C and 2 min at 72°C, with a final extension step at 72°C for 10 min. For the nifH primers, an initial denaturation step at 94°C for 5 min and a final elongation step of 10 min at 72°C were added to the original programme (Zehr & McReynolds, 1989). The hetR amplification was run according to Janson et al. (1998). Negative (without any DNA template) and positive control reactions were carried out simultaneously. PCR products were checked by electrophoresis on 1.5% agarose gels at 80 V for 1 h 30 min, followed by ethidium bromide staining and visualization on an UV transilluminator.
Sequencing, nucleotide sequence accession numbers and sequence comparative analysis
Purification of amplicons was performed with the Jetquick PCR Product Purification Spin Kit (Genomed, Löhne, Germany) before sequencing (STABVida, Portugal). A BLAST search was used to detect similar sequences deposited in the GenBank™ database of NCBI (http://www.ncbi.nlm.nih.gov). ClustalX ver. 1.83 (Thompson et al., 1997) was used to align sequences and reconstruct trees using the neighbour-joining (NJ) method (with bootstrap values from 1,000 NJ bootstrap replicates). Phylogenetic analyses of sequences were carried out using PAUP* ver. 4.0b10 (Swofford, 2003) for maximum-parsimony analyses and MrBayes ver. 3.0b4 (Ronquist & Huelsenbeck, 2003) for Bayesian analyses. Maximum-parsimony analyses were performed using the heuristic search option with 1,000 random taxa additions and three bisections and reconnections as the branch-swapping algorithm. All characters were unordered and of equal weight, and alignment gaps were treated as missing data. Branches of zero length were collapsed and all multiple, equally parsimonious trees were saved. The robustness of the most parsimonious trees was evaluated by 1,000 bootstraps replications. Bayesian analyses employing a Markov chain Monte Carlo (MCMC) method were performed. The general time-reversible model of evolution (Rodríguez et al., 1990) was used, including estimation of invariable sites and assuming a discrete gamma distribution with six rate categories (GTR+Γ+G). Four MCMC chains were run simultaneously, starting from random trees for 1,000,000 generations. Trees were sampled every 100th generation for a total of 10,000 trees. The first 1,000 trees were discarded as the burn-in phase of each analysis. Posterior probabilities (Rannala & Yang, 1996) were determined from a majority-rule consensus tree generated with the remaining 9,000 trees. Visualization of trees was performed by using the TreeView 1.6.6 programme (Page, 1996). The cyanobacterial partial sequences determined in the present study were deposited in the GenBank™ database under the accession numbers GQ443447 to GQ443452.
Statistical treatment
Statistical differences related to fatty acid analysis were obtained by using one-way ANOVA with the nonparametric Tukey’s post-test. A value of P < 0.05 was considered statistically significant. Data were analysed using GraphPad Prism 5.01 (GraphPad Software, Inc., San Diego, CA, USA). In order to confirm the reproducibility of the results, all data related to fatty acid and morphometric analyses were expressed as mean ± standard error (SE) of at least 3 and 50 independent observations, respectively.
Results
Morphological characterization of strains
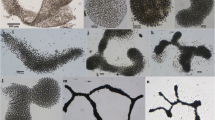
The morphometric characteristics of the three strains are shown in Table 1. By comparative analysis with the available literature, the strain UTAD_A212 was identified as Anabaena cylindrica Lemm. (Fig. 1a). This species has straight and solitary trichomes, with neither branching nor tapering; vegetative cells are cylindrical, without aerotopes and apical cells are rounded; heterocysts are long rounded and mostly intercalary and single; colourless akinetes are cylindrical, close to the heterocyst and have a smooth wall. The strain UTAD_N213 was identified as Nostoc muscorum C. Agardh (Fig. 1, b1 and b2). It is characterized by thickly entangled straight trichomes with neither branching nor tapering; vegetative cells are short barrel-shaped or cylindrical, without aerotopes and apical cells are rounded; heterocysts are single, subspherical and intercalary; when visible, akinetes are oblong with a yellow smooth wall. The morphometry data corroborate the identification of the strain UADFA16 as Aphanizomenon gracile Lemm. (Fig. 1c) (de Figueiredo et al., 2010). It has free-floating, straight and solitary trichomes narrowed towards the ends; vegetative cells are barrel-shaped, with aerotopes and apical cells are rounded; heterocysts are solitary, mostly short and cylindrical, sometimes broadly oval to nearly spherical, and typically wider than vegetative cells; akinetes are cylindrical, usually solitary and distant from heterocysts.

Photomicrographs of the cyanobacterial isolates: a Anabaena cylindrica UTAD_A212; b1, b2 Nostoc muscorum UTAD_N213; c Aphanizomenon gracile UADFA16. In addition to vegetative cells, heterocysts (het) and hormogonia (hor) are seen. The small inset in each photomicrograph shows the akinete germination pattern of each strain. Scale bar corresponds to 5 μm
Fatty acids analysis
The FAME composition of the total lipid extracts of cyanobacterial strains is shown in Fig. 2. The predominant cellular FAMEs in lipids of the three cyanobacterial strains were palmitic (16:0), oleic (18:1ω9) and α-linolenic (18:3ω3). Additionally, palmitoleic (16:1), stearic (18:0) and linoleic (18:2ω6) acids were present. They presented trace amounts of myristic acid (14:0) and C20 polyunsaturated fatty acids. Figure 2 shows the significantly higher content of palmitic acid of UADFA16 (48.18% of the total fatty acids; TFAs) compared to UTAD_A212 (22.51% of TFAs) and UTAD_N213 (16.92% of TFAs). The content of oleic acid was also significantly high in UADFA16 (22.12%), followed by UTAD_A212 and UTAD_N213, with 13.95 and 9.73% of TFAs, respectively. Strains UTAD_A212 and UTAD_N213 contained higher concentrations of α-linolenic acid than UADFA16 (4.78%), with values relative to TFAs of 23.13 and 17.19%, respectively. The content of stearic acid was significantly different in the three strains, with 12.82, 8.93 and 2.72% for UTAD_A212, UTAD_N213 and UADFA16, respectively. The lowest linoleic acid content was recorded in UADFA16 (2.12%), followed by UTAD_A212 (5.45%) and UTAD_N213 (9.32%). The FAME distribution pattern of palmitoleic acid was similar to that of linoleic acid among the three strains, with 2.67, 3.76 and 5.87% for UADFA16, UTAD_A212 and UTAD_N213, respectively.

Fatty acid composition of the cyanobacterial strains. The values were expressed in % of total fatty acids and are means ± SE of at least three independent experiments. Results of the one-way ANOVA factorial analysis. Values with a common letter are not significantly different according to Tukey’s test (P < 0.05)
The saturated fatty acids (SAFAs) in UADFA16 accounted for 61.74% of the TFAs, being significantly higher than the SAFA content in UTAD_A212 (46.27%) and UTAD_N213 (45.76%) (Table 2). Strain UADFA16 also ranked first in terms of monounsaturated fatty acids (MUFAs) with 24.78% of TFAs, followed by UTAD_A212 (17.71%) and UTAD_N213 (15.60%). UTAD_N213 had the highest polyunsaturated fatty acids (PUFAs) content (38.64%), followed by UTAD_A212 and UADFA16, with 36.01 and 13.48%, respectively. The proportion of SAFA to unsaturated fatty acids in UADFA16 was approximately twice as high as UTAD_A212 and UTAD_N213, whereas the ratio MUFA:PUFA in UADFA16 was 3.8 and 4.5 times higher than in UTAD_A212 and UTAD_N213, respectively. However, the unsaturation index (UI) was significantly high in UTAD_N213 (142.78), followed by UTAD_A212 (131.93) and UADFA16 (73.21).
Partial sequencing of 16S rRNA, hetR and nifH genes from cyanobacterial strains
The strain UADFA16 had high 16S rDNA sequence similarity (99%) with Aphanizomenon gracile 219 (isolated from a Danish lake) and Anabaena flos-aquae PCC 7905 (type strain) (Fig. 3). The strain UTAD_A212 had 99% similarity with a 16S rDNA sequence of Anabaena cylindrica NIES19, also named Anabaena cylindrica PCC 7122 and isolated from Japan, while strain UTAD_N213 showed 98% sequence similarity with Nostoc spp. such as Nostoc sp. 8938 or Nostoc muscorum I. According to Fig. 4 and the hetR gene fragments (with approximately 450 bp), strain UADFA16 showed 98% similarity with Anabaena flos-aquae SAG 30.87 and 97% similarity Aphanizomenon sp. TR183, whereas strain UTAD_N213 showed a total match with Nostoc PCC 7906; however, strain UTAD_A212 showed similarity values of 86% (for Cylindrospermum sp. A1345) or below. The sequence of the nifH gene fragment (319 bp long) of strain UADFA16 had high similarity (98%) with a nitrogenase reductase sequence of an uncultured cyanobacterium clone Gt1463 (isolated from the Baltic Sea, Sweden) (Fig. 5). Strain UTAD_N213 nifH partial sequence showed high similarity (99%) with Nostoc muscorum CC1090A1 followed by an uncultured soil bacterium and Nostoc commune UTEX 584, with 97 and 96% similarities, respectively. For strain UTAD_A212, similarities of 99 and 100% were found with the partial nitrogenase reductase gene sequences of a freshwater strain of Anabaena oscillarioides and Anabaena cylindrica UTEX B629, respectively.

Maximum-parsimony phylogenetic tree constructed from 16S rRNA gene partial sequences (670 bp) from cultured cyanobacterial isolates (sequences with accession numbers). Synechococcus sp. PS845 (AF448070) sequence was used as outgroup. Percentages of 1,000 bootstraps replicates are indicated near the nodes (only those values >50% are shown) following the order maximum-parsimony/posterior probability/neighbour-joining (NJ) algorithm. The scale bar represents 10 changes

Maximum-parsimony phylogenetic tree constructed from hetR gene partial sequences (approximately 450 bp) from cultured cyanobacterial isolates (sequences with accession numbers). A sequence from Trichodesmium pelagicum were used as outgroup. Percentages of 1,000 bootstraps replicates are indicated near the nodes (only those values >50% are shown) following the order maximum-parsimony/posterior probability/neighbour-joining (NJ) algorithm. The scale bar represents 10 changes

Maximum-parsimony phylogenetic tree constructed from nifH gene partial sequences (approximately 330 bp) from cultured cyanobacterial isolates (sequences with accession numbers). The sequence from a Synechococcus sp. (SSU22146) was used as outgroup. Percentages of 1,000 bootstraps replicates are indicated near the nodes (only those values >50% are shown) following the order maximum-parsimony/posterior probability/neighbour-joining (NJ) algorithm.The scale bar represents 10 changes
Discussion
The classification of cyanobacteria has routinely relied on morphological characteristics which are not always trustworthy, as they may show variation depending on culturing and environmental conditions (Nayak et al., 2007), and lead to misidentifications (Komárek & Anagnostidis, 1989). These problems of traditional morphological classification, together with the lack of molecular data, pose serious hindrances for taxonomy and systematics of cyanobacteria (Hayes et al., 2007; Komárek, 2010). Notwithstanding, the morphological assignment of some species belonging to the genera Anabaena, Aphanizomenon and Nostoc has been discussed recently by some researchers (Hindák, 2000; Lyra et al., 2001; Rajaniemi et al., 2005; Komárek & Komárková, 2006; Zapomělová et al., 2010). According to morphological and molecular data, the genus Anabaena has been recently reviewed and it includes now only the species without gas vesicles, whereas the new genus Dolichospermum includes the former planktonic Anabaena species (which are closely related to the Aphanizomenon genus) (Komárek, 2006; Wacklin et al., 2009; Komárek, 2010). The morphological distinction between some species of the genera Dolichospermum and Aphanizomenon is problematic (Rajaniemi et al., 2005; de Figueiredo et al., 2010). The same happens between the genera Anabaena/Dolichospermum and Nostoc, whose identification traditionally relied on slime production (Geitler, 1932), the nature and shape of filaments, shape of end cells (Lachance, 1981), and the pattern of akinete formation/position of akinete towards heterocyst (Desikachary, 1959). Molecular-based studies have also supported their distinction as different genera (Henson et al., 2002; Rajaniemi et al., 2005). In the present study, morphological characteristics of the studied strains (belonging to the family Nostocaceae) revealed remarkable morphological differences between them and were congruent with recent studies but also with traditional references.
Besides morphological examination, it is currently accepted that characterization and taxonomy of cyanobacteria must combine multidisciplinary approaches (Castenholz & Waterbury, 1989; Wilmotte, 1994; Gillis et al., 2005; Hayes et al., 2007; Komárek, 2010). This so-called polyphasic methodology (including phenotypic, chemotaxonomic and genotypic data) has been increasingly followed by many cyanobacteriologists worldwide, e.g., Rajaniemi et al. (2005), Willame et al. (2006), Nayak et al. (2007), Li et al. (2008), Saker et al. (2009) and Schleifer (2009). Among the molecular methods, the analysis of the 16S rRNA gene sequences has proved to be a useful tool for exploring phylogenetic relationships among cyanobacteria (Gugger et al., 2002a; Rajaniemi et al., 2005; Willame et al., 2006; Li et al., 2008; Pan et al., 2008; Han et al., 2009; Saker et al., 2009; Komárek, 2010; Zapomělová et al., 2010). However, it is important to highlight that, although the 16S rRNA gene contains variable regions, it is so conserved that it may not always provide a good resolution at the species or intra-species levels in some cyanobacterial groups (Wilmotte, 1994; Lyra et al., 2001; Li & Watanabe, 2004). The unique distribution of the hetR gene among filamentous cyanobacteria and its relatively high sequence variation between closely related strains makes it also a powerful gene marker for discriminating filamentous cyanobacteria (Janson et al., 1998; Han et al., 2009). Moreover, the nifH gene encoding the iron protein subunit of nitrogenase is highly conserved among all diazotrophs and its phylogeny has been shown to resemble largely the 16S rRNA phylogeny (Zehr & McReynolds, 1989; Wartiainen et al., 2008). The phylogenetic analyses of the partially sequenced genes (16S rRNA, hetR and nifH) allowed the discrimination of the three studied strains in distinct clusters belonging to the genera Anabaena, Aphanizomenon and Nostoc. Nevertheless, at the species level, there are hindrances due to the lack of sequences deposited so far concerning the species of the studied strains, particularly N. muscorum and An. cylindrica. For example, the low similarity percentage match obtained for the hetR sequence of An. cylindrica UTAD_A212 is due to the lack of deposited hetR sequences so far concerning the genus Anabaena and, particularly, the species An. cylindrica. This highlights the need for more deposited sequences of this genus/species and the importance of the data made available in the present study for future phylogenetic analyses.
Characteristic macromolecules of the cell such as fatty acids, isozymes and other proteins have also been used in taxonomic studies of cyanobacterial strains (Wilmotte, 1994; Saker et al., 2009). The composition of whole-cell fatty acid as chemotaxonomic marker has been extensively used for cyanobacterial taxonomic purposes. The method is cheap, rapid and efficient, and allows cyanobacterial differentiation at the levels of genus and species (Kenyon et al., 1972; Wilmotte, 1994; Gugger et al., 2002b; Gillis et al., 2005; Nayak et al., 2007; Temina et al., 2007; Li et al., 2008). Besides, fatty acids are primary metabolites of the acetyl CoA pathway, which is genetically determined, evolutionarily very old and therefore conservative (Petkov & Garcia, 2007). For this reason, fatty acid composition and the relative percentage of individual components in each genus or species can reflect evolutionary changes as well as metabolic and biochemical processes in cyanobacteria (Caudales & Wells, 1992). Generally, the obtained results clearly showed significant differences in the qualitative and quantitative variations of FAMEs in the three strains. The main identified FAMEs were in accordance with those previously found in the genera Anabaena, Aphanizomenon and Nostoc (Caudales & Wells, 1992; Wada & Murata, 1998; Li & Watanabe, 2001, 2004; Gugger et al., 2002a, b; Temina et al., 2007). Generally, the major FAMEs in cyanobacteria are palmitic, palmitoleic, hexadecadienoic, stearic and oleic, and differences in their composition have led to classification of cyanobacteria into four groups (Kenyon et al., 1972; Murata et al., 1992): organisms of the first group are devoid of polyunsaturated fatty acids (PUFA) and contain only saturated (SAFA) plus mono-unsaturated (MUFA) fatty acids; the second and third groups contain either α-linolenic acid (18:3ω3) or γ-linolenic acid (18:3ω6), respectively and the fourth group contains also octadecatetraenoic acid (18:4ω3). Therefore, according to the Kenyon–Murata classification system, the fatty acid composition of the three strains corresponds to Group II.
Dolichospermum strains have been differentiated based on their FAME composition and a correlation with morphological characteristics has been reported (Li & Watanabe, 2001, 2004). Nevertheless, Anabaena and Aphanizomenon strains that appear morphologically distinct have shown a high degree of similarity in cellular FAME profiles (Gugger et al., 2002b), as well as in their ribulose-1,5-bisphosphate carboxylase/oxygenase composition and 16S rDNA sequences (Gugger et al., 2002a). In our study, the strains that belong to Anabaena and Aphanizomenon genera had significantly different FAME patterns. However, it must be noticed that the small number of investigated strains belonging to these genera hinders clear taxonomic conclusions at the generic level, thus emphasizing the need for further studies. Interestingly, the percentages of total saturated fatty acids of An. cylindrica UTAD_A212 and N. muscorum UTAD_N213 were not significantly different between them and were within the range previously obtained in Nostoc isolates (Caudales & Wells, 1992). This suggests that Anabaena and Nostoc genera may not be differentiated by the saturated to unsaturated fatty acid ratio, as defended by Caudales & Wells (1992). Horizontal gene transfer may be an important factor for inter-genera similarity of fatty acids composition, as previously reported for cyanobacteria (Thacker & Paul, 2004; Welker & von Döhren, 2006). Moreover, similar environmental conditions from collection locations may also trigger similar chemical responses (Thacker & Paul, 2004). N. muscorum is a terrestrial species in origin, but may also be distributed in very varied environments (see e.g., Oudra et al., 2009). In rice fields, this strain can be subjected to dryness and severe changes of other abiotic factors, such as temperature and light intensity. In response to changes and fluctuations in these environmental factors, it shows a high proportion of unsaturated fatty acids (Wada & Murata, 1998), leading to a high fatty acid unsaturation index. The high PUFA content of N. muscorum relatively to other species has been reported (Liu et al., 2003). Caudales & Wells (1992) also found that strains of Anabaena and Nostoc genera were high in PUFAs and have MUFA:PUFA ratios averaging less than 1.0. These results are congruent with those obtained in our study. On the other hand, the high MUFA:PUFA ratio of Aph. gracile UADFA16 clearly separated this strain from the other two and contributed to the lowest UI value obtained, which was undoubtedly a consequence of the extremely high percentage of palmitic acid detected in this strain, also confirmed by GC–MS (data not shown). Accordingly, our results suggested that the MUFA:PUFA ratio, together with the UI, could be used as genus-specific chemotaxonomic biomarkers in the differentiation of Nostocaceae members. However, Temina et al. (2007) suggested that, for FAME composition, the ecosystem from which cyanobacteria were collected is more important than their taxonomic affinity. This evidence, as well as potential horizontal gene transfer, may explain the similarity found between the FAME patterns of An. cylindrica and N. muscorum strains, both isolated from rice fields, which were very distinct from the FAME patterns of the planktonic Aph. gracile strain isolated from Vela Lake. This latter species is known to be planktonic in other Portuguese water bodies with high water retention times (Pereira et al., 2004). Besides Portuguese rice fields, An. cylindrica was also found in different benthic habitats worldwide (Rajaniemi et al., 2005; Pan et al., 2008). The genus Nostoc is also predominant in rice fields worldwide, as in Portugal (Galhano et al., 2009), and has a beneficial role as a natural biofertilizer in increasing soil fertility (Whitton, 2000). However, the potential harmful toxicity of N. muscorum UTAD_N213 must be checked in future studies since another similar strain was recently found dominant in a toxic bloom in the Oukaïmeden river, near Marrakech (Morocco) (Oudra et al., 2009). In effect, these authors found the existence of a benthic microcystin-producing N. muscorum strain under the particular environmental conditions of the Oukaïmeden region, which may be a potential human health hazard. Notwithstanding this, the high proportion of palmitic acid obtained in Aph. gracile UADFA16 is very interesting and deserves further biochemical and molecular investigations. This strain could be a potential source of commercially interesting fatty acids. For the same reason, the relative high percentage of the essential FAME α-linolenic acid of Anabaena and Nostoc strains must also be taken in account.
Conclusions
A polyphasic methodology, which integrates phenotypic, chemotaxonomic and genotypic data, is undoubtedly becoming imperative for a better characterization of cyanobacterial strains. In the present study, results from morphometric and genetic criteria showed congruency, in spite of the lack of sequences deposited to date concerning the species N. muscorum and An. cylindrica. Together with morphological and genotypic analysis, fatty acid characterization proved to be a valuable chemotaxonomic tool, as shown by the differentiation between the genera Anabaena, Aphanizomenon and Nostoc. However, the results suggest that environmental stresses must be taken into account since they might have a major role in the production of unsaturated fatty acids. The high proportion of palmitic acid obtained in Aph. gracile UADFA16 was rather interesting and should be further explored both in biochemical and molecular investigations, since this strain could be a potential source of commercially interesting fatty acids. This study contains important contributions for further research on: (1) the beneficial exploration of An. cylindrica and N. muscorum strains as potential biofertilizers in Portuguese rice fields exposed to local applied agrochemicals; and (2) the physiological, cellular, biochemical and molecular responses of the studied strains to environmental changes in the studied and other freshwater ecosystems. Further research to investigate horizontal gene transfer, the inclusion of more strains representative of each habitat, and a deeper screening of more chemotaxonomic and molecular biomarkers will be advisable in future studies.
References
Andersen, R. A. & M. Kawachi, 2005. Traditional microalgae isolation techniques. In Andersen, R. A. (ed.), Algal Culturing Techniques. Elsevier Academic Press, Tokyo: 83–100.
Ariosa, Y., D. Carrasco, A. Quesada & E. Fernandez-Valiente, 2006. Incorporation of different N sources and light response curves of nitrogenase and photosynthesis by cyanobacterial blooms from rice fields. Microbial Ecology 51: 394–403.
Castenholz, R. W., 1989. Subsection IV. Order Nostocales. In Staley, J. T., M. P. Bryant, N. Pfennig & J. G. Holt (eds), Bergey’s Manual of Systematic Bacteriology, Vol. 3. Williams & Wilkins, Baltimore: 1780–1793.
Castenholz, R. W. & J. B. Waterbury, 1989. Group I. Cyanobacteria. In Staley, J. T., M. P. Bryant, N. Pfennig & J. G. Holt (eds), Bergey’s Manual of Systematic Bacteriology, Vol. 3. Williams & Wilkins, Baltimore: 1710–1727.
Caudales, R. & J. M. Wells, 1992. Differentiation of the free-living Anabaena and Nostoc cyanobacteria on the basis of fatty acid composition. International Journal of Systematic and Evolutionary Microbiology 42: 246–251.
de Figueiredo, D. R., A. S. S. P. Reboleira, S. C. Antunes, N. Abrantes, U. Azeiteiro, F. Gonçalves & M. J. Pereira, 2006. The effect of environmental parameters and cyanobacterial blooms on phytoplankton dynamics of a Portuguese temperate lake. Hydrobiologia 568: 145–157.
de Figueiredo, D. R., A. Alves, M. J. Pereira & A. Correia, 2010. Molecular characterization of bloom-forming Aphanizomenon strains isolated from Vela Lake (Western Central Portugal). Journal of Plankton Research 32: 239–252.
Desikachary, T. V., 1959. Cyanophyta. Indian Council of Agricultural Research, New Delhi, India.
Galhano, V., F. Peixoto, J. Gomes-Laranjo & E. Fernández-Valiente, 2009. Differential effects of bentazon and molinate on Anabaena cylindrica, an autochthonous cyanobacterium of Portuguese rice field agro-ecosystems. Water, Air and Soil Pollution 197: 211–222.
Geitler, L., 1932. Cyanophyceae. In Kolkwitz, R. (ed.), Rabenhorst’s Kriptogamenflora von Deutschland, Österreich und der Schweiz. Akademische Verlagsgesellschaft, Leipzig, Federal Republic of Germany: 1–1196.
Gillis, M., P. Vandamme, P. De Vos, J. Swings & K. Kersters, 2005. Polyphasic taxonomy. In Brenner, D. J., N. R. Krieg, J. T. Staley & G. M. Garrity (eds), Bergey’s Manual of Systematic Bacteriology. Springer, New York: 43–48.
Gugger, M., C. Lyra, P. Henriksen, A. Couté, J.-F. Humbert & K. Sivonen, 2002a. Phylogenetic comparison of the cyanobacterial genera Anabaena and Aphanizomenon. International Journal of Systematic and Evolutionary Microbiology 52: 1867–1880.
Gugger, M., C. Lyra, I. Suominen, I. Tsitko, J.-F. Humbert, M. S. Salkinoja-Salonen & K. Sivonen, 2002b. Cellular fatty acids as chemotaxonomic markers of the genera Anabaena, Aphanizomenon, Microcystis, Nostoc and Planktothrix (cyanobacteria). International Journal of Systematic and Evolutionary Microbiology 52: 1007–1015.
Guillard, R. R. L., 2005. Purification methods for microalgae. In Andersen, R. A. (ed.), Algal Culturing Techniques. Elsevier Academic Press, Tokyo: 117–132.
Han, D., Y. Fan & Z. Hu, 2009. An evaluation of four phylogenetic markers in Nostoc: Implications for cyanobacterial phylogenetic studies at the intrageneric level. Current Microbiology 58: 170–176.
Hayes, P. K., N. A. El Semary & P. Sánchez-Baracaldo, 2007. The taxonomy of cyanobacteria: molecular insights into a difficult problem. In Brodie, J. & J. Lewis (eds), Unravelling The Algae: The Past, Present, and Future of Algal Systematics. CRC Press/Taylor & Francis Group, Boca Raton: 93–101.
Henson, B. J., L. E. Watson & S. R. Barnum, 2002. Molecular differentiation of the heterocystous cyanobacteria, Nostoc and Anabaena, based on complete NifD sequences. Current Microbiology 45: 161–164.
Hindák, F., 2000. Morphological variation of four planktic nostocalean cyanophytes – members of the genus Aphanizomenon or Anabaena? Hydrobiologia 438: 107–116.
Janson, S., A. Matveyev & B. Bergman, 1998. The presence and expression of hetR in the non-heterocystous cyanobacterium Symploca PCC 8002. FEMS Microbiology Letters 168: 173–179.
Kenyon, C. N., R. Rippka & R. Y. Stanier, 1972. Fatty acid composition and physiological properties of some filamentous blue-green algae. Archives of Microbiology 83: 216–236.
Komárek, J., 2006. Cyanobacterial taxonomy: current problems and prospects for the integration of traditional and molecular approaches. Algae 21: 349–375.
Komárek, J., 2010. Modern taxonomic revision of planktic nostocacean cyanobacteria: a short review of genera. Hydrobiologia 639: 231–243.
Komárek, J. & K. Anagnostidis, 1989. Modern approach to the classification system of Cyanophytes. 4: Nostocales. Archive für Hydrobiologie Supplement/Algological Studies 82(3/56): 247–345.
Komárek, J. & J. Komárková, 2006. Diversity of Aphanizomenon-like cyanobacteria. Czech Phycology, Olomouc 6: 1–32.
Lachance, M., 1981. Genetic relatedness of heterocystous cyanobacteria by deoxyribonucleic acid-deoxyribonucleic acid reassociation. International Journal of Systematic and Evolutionary Microbiology 31: 139–147.
Lane, D. J., 1991. 16S/23S rRNA sequencing. In Stackebrandt, E. & M. Goodfellow (eds), Nucleic Acid Techniques in Bacterial Systematics. Wiley, Chichester, UK: 115–175.
Li, R. & M. M. Watanabe, 2001. Fatty acid profiles and their chemotaxonomy in planktonic species of Anabaena (Cyanobacteria) with straight trichomes. Phytochemistry 57: 727–731.
Li, R. & M. M. Watanabe, 2004. Fatty acid composition of planktonic species of Anabaena (Cyanobacteria) with coiled trichomes exhibited a significant taxonomic value. Current Microbiology 49: 376–380.
Li, R., S. W. Wilhelm, W. W. Carmichael & M. M. Watanabe, 2008. Polyphasic characterization of water bloom forming Raphidiopsis species (cyanobacteria) from central China. Harmful Algae 7: 146–153.
Liu, X.-J., F. Chen & Y. Jiang, 2003. Differentiation of Nostoc flagelliforme and its neighboring species using fatty-acid profiling as a chemotaxonomic tool. Current Microbiology 47: 467–474.
Lyra, C., S. Suomalainen, M. Gugger, C. Vezie, P. Sundman, L. Paulin & K. Sivonen, 2001. Molecular characterization of planktic cyanobacteria of Anabaena, Aphanizomenon, Microcystis and Planktothrix genera. International Journal of Systematic and Evolutionary Microbiology 51: 513–526.
Marques, C. R., N. Abrantes, D. R. de Figueiredo, M. J. Pereira & F. Gonçalves, 2008. Are Pseudokirchneriella subcapitata and Chlorella vulgaris affected by environmental samples from a rice field? Water, Air and Soil Pollution 189: 49–59.
Murata, N., H. Wada & Z. Gombos, 1992. Modes of fatty-acid desaturation in cyanobacteria. Plant Cell Physiology 33: 933–941.
Nayak, S., R. Prasanna, B. M. Prasanna & D. B. Sahoo, 2007. Analysing diversity among Indian isolates of Anabaena (Nostocales, Cyanophyta) using morphological, physiological and biochemical characters. World Journal of Microbiology and Biotechnology 23: 1575–1584.
Nübel, U., F. Garcia-Pichel & G. Muyzer, 1997. PCR primers to amplify 16S rRNA genes from cyanobacteria. Applied and Environmental Microbiology 63: 3327–3332.
Oudra, B., M. D. E. Andaloussi & V. M. Vasconcelos, 2009. Identification and quantification of microcystins from a Nostoc muscorum bloom occurring in Ouka meden River (High-Atlas mountains of Marrakech, Morocco). Environmental Monitoring and Assessment 149: 437–444.
Page, R. D. M., 1996. TreeView: an application to display phylogenetic trees on personal computers. Computer Applications in the Biosciences 12: 357–358.
Pan, X. J., F. Y. Chang, L. J. Kang, G. B. Li, D. H. Li, Y. D. Liu, Y. W. Shen & Z. H. Wei, 2008. Morphological characteristics and phylogenetic relationship of Anabaena species from Lakes Dianchi and Erhai, China. Hydrobiologia 614: 353–362.
Pereira, P., R. H. Li, W. W. Carmichael, E. Dias & S. Franca, 2004. Taxonomy and production of paralytic shellfish toxins by the freshwater cyanobacterium Aphanizomenon gracile LMECYA40. European Journal of Phycology 39: 361–368.
Petkov, G. & G. Garcia, 2007. Which are fatty acids of the green alga Chlorella? Biochemical Systematics and Ecology 35: 281–285.
Quesada, A., E. Moreno, D. Carrasco, T. Paniagua, L. Wormer, C. De Hoyos & A. Sukenik, 2006. Toxicity of Aphanizomenon ovalisporum (Cyanobacteria) in a Spanish water reservoir. European Journal of Phycology 41: 39–45.
Rajaniemi, P., P. Hrouzek, K. Kaštovská, R. Willame, A. Rantala, L. Hoffmann, J. Komárek & K. Sivonen, 2005. Phylogenetic and morphological evaluation of the genera Anabaena, Aphanizomenon, Trichormus and Nostoc (Nostocales, Cyanobacteria). International Journal of Systematic and Evolutionary Microbiology 55: 11–26.
Rannala, B. & Z. H. Yang, 1996. Probability distribution of molecular evolutionary trees: a new method of phylogenetic inference. Journal of Molecular Evolution 43: 304–311.
Rasmussen, B., I. R. Fletcher, J. J. Brocks & M. R. Kilburn, 2008. Reassessing the first appearance of eukaryotes and cyanobacteria. Nature 455: 1101–1104.
Rastogi, R. P. & R. P. Sinha, 2009. Biotechnological and industrial significance of cyanobacterial secondary metabolites. Biotechnology Advances 27: 521–539.
Rippka, R., J. Deruelles, J. B. Waterbury, M. Herdman & R. Y. Stanier, 1979. Generic assignments, strain histories and properties of pure cultures of cyanobacteria. Journal of General Microbiology 111: 1–61.
Rodríguez, F., J. L. Oliver, A. Marín & J. R. Medina, 1990. The general stochastic model of nucleotide substitutions. Journal of Theoretical Biology 142: 485–501.
Ronquist, F. & J. P. Huelsenbeck, 2003. MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics 19: 1572–1574.
Rosa, D. & A. Catalá, 1998. Fatty acid profiles and non enzymatic lipid peroxidation of microsomes and mitochondria from bovine liver, kidney, lung and heart. Archives of Physiology and Biochemistry 106: 33–37.
Rücker, J., A. Stüken, B. Nixdorf, J. Fastner, I. Chorus & C. Wiedner, 2007. Concentrations of particulate and dissolved cylindrospermopsin in 21 Aphanizomenon-dominated temperate lakes. Toxicon 50: 800–809.
Saker, M., C. Moreira, J. Martins, B. Neilan & V. M. Vasconcelos, 2009. DNA profiling of complex bacterial populations: toxic cyanobacterial blooms. Applied Microbiology and Biotechnology 85: 237–252.
Sato, N. & N. Murata, 1988. Membrane lipids. Methods in Enzymology 167: 251–259.
Schaeffer, D. J. & V. S. Krylov, 2000. Anti-HIV activity of extracts and compounds from algae and cyanobacteria. Ecotoxicology and Environmental Safety 45: 208–227.
Schleifer, K. H., 2009. Classification of Bacteria and Archaea: past, present and future. Systematic and Applied Microbiology 32: 533–542.
Stüken, A., R. J. Campbell, A. Quesada, A. Sukenik, P. K. Dadheech & C. Wiedner, 2009. Genetic and morphologic characterization of four putative cylindrospermopsin producing species of the cyanobacterial genera Anabaena and Aphanizomenon. Journal of Plankton Research 31: 465–480.
Swofford, D. L., 2003. PAUP*. Phylogenetic Analysis Using Parsimony (and other Methods), Version 4.0b10 [Computer Program CD-ROM]. Sinauer Associates, Sunderland, MA.
Temina, M., H. Rezankova, T. Rezanka & V. M. Dembitsky, 2007. Diversity of the fatty acids of the Nostoc species and their statistical analysis. Microbiological Research 162: 308–321.
Thacker, R. W. & V. J. Paul, 2004. Morphological, chemical, and genetic diversity of tropical marine cyanobacteria Lyngbya spp. and Symploca spp. (Oscillatoriales). Applied and Environmental Microbiology 70: 3305–3312.
Thompson, J. D., T. J. Gibson, F. Plewniak, F. Jeanmougin & D. G. Higgins, 1997. The CLUSTAL_X windows interface: flexible strategies for multiple sequence alignment aided by quality analysis tools. Nucleic Acids Research 25: 4876–4882.
Valério, E., P. Pereira, M. L. Saker, S. Franca & R. Tenreiro, 2005. Molecular characterization of Cylindrospermopsis raciborskii strains isolated from Portuguese freshwaters. Harmful Algae 4: 1044–1052.
Wacklin, P., L. Hoffmann & J. Komárek, 2009. Nomenclatural validation of the genetically revised cyanobacterial genus Dolichospermum (RALFS ex BORNET et FLAHAULT) comb. nova. Fottea 9: 59–64.
Wada, H. & N. Murata, 1998. Membrane lipids in cyanobacteria. In Siegenthaler, P.-A. & N. Murata (eds), Lipids in Photosynthesis: Structure, Function and Genetics. Kluwer Academic Publishers, Dordrecht: 65–81.
Wartiainen, I., T. Eriksson, W. Zheng & U. Rasmussen, 2008. Variation in the active diazotrophic community in rice paddy: nifH PCR-DGGE analysis of rhizosphere and bulk soil. Applied Soil Ecology 39: 65–75.
Welker, M. & H. von Döhren, 2006. Cyanobacterial peptides – nature’s own combinatorial biosynthesis. FEMS Microbiology Reviews 30: 530–563.
Whitton, B. A., 2000. Soils and rice-fields. In Whitton, B. A. & M. Potts (eds), The Ecology of Cyanobacteria: Their Diversity in Time and Space. Kluwer Academic Publishers, Dordrecht: 233–255.
Willame, R., C. Boutte, S. Grubisic, A. Wilmotte, J. Komárek & L. Hoffmann, 2006. Morphological and molecular characterization of planktonic cyanobacteria from Belgium and Luxembourg. Journal of Phycology 42: 1312–1332.
Wilmotte, A., 1994. Molecular evolution and taxonomy of the cyanobacteria. In Bryant, D. A. (ed.), The Molecular Biology of Cyanobacteria. Kluwer Academic Publishers, Dordrecht: 1–25.
Zancan, S., R. Trevisan & M. G. Paoletti, 2006. Soil algae composition under different agro-ecosystems in North-Eastern Italy. Agriculture Ecosystems and Environment 112: 1–12.
Zapomělová, E., K. Řehaková, J. Jezberová & J. Komárková, 2010. Polyphasic characterization of eight planktonic Anabaena strains (Cyanobacteria) with reference to the variability of 61 Anabaena populations observed in the field. Hydrobiologia 639: 99–113.
Zehr, J. P. & L. A. McReynolds, 1989. Use of degenerate oligonucleotides for amplification of the nifH gene from the marine cyanobacterium Trichodesmium thiebautii. Applied and Environmental Microbiology 55: 2522–2526.
Zwart, G., M. P. Kamst-van Agterveld, I. van der Werff-Staverman, F. Hagen, H. L. Hoogveld & H. J. Gons, 2005. Molecular characterization of cyanobacterial diversity in a shallow eutrophic lake. Environmental Microbiology 7: 365–377.
Acknowledgments
This study was supported by PhD grants to Victor Galhano (SFRH/BD/17582/2004) and Daniela de Figueiredo (SFRH/BD/23864/2005) of Portuguese Science and Technology Foundation (FCT), Ministry of Science, Technology and Higher Education, Portuguese Government. The authors are indebted to Prof. Fernando Nunes (Chemistry Department, UTAD, Vila Real) for instrumental laboratory facilities on lipid analysis and to Dr. Fátima Santos (Botany Department, FCTUC, Coimbra) for her kind help on morphological identification and loan of precious historical publications. Further, we are thankful to Prof. Luigi Naselli-Flores (Department of Botanical Sciences, University of Palermo, Palermo, Italy) and to the anonymous reviewer for their constructive and helpful comments on the manuscript.
Author information
Authors and Affiliations
Corresponding author
Additional information
Handling editor: Luigi Naselli-Flores
Rights and permissions
About this article
Cite this article
Galhano, V., de Figueiredo, D.R., Alves, A. et al. Morphological, biochemical and molecular characterization of Anabaena, Aphanizomenon and Nostoc strains (Cyanobacteria, Nostocales) isolated from Portuguese freshwater habitats. Hydrobiologia 663, 187–203 (2011). https://doi.org/10.1007/s10750-010-0572-5
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10750-010-0572-5