
Abstract
Nectar-related traits influence the number of bee visits to sunflowers, but the effects of floret size on the diversity and composition of bee communities are unknown. In 4 year × planting date combinations, bees were collected from sunflowers with small (< 7.5 mm), medium (7.5–9 mm), or large (> 9 mm) florets, and counts of foraging bees were used to clarify bee responses to plant traits. No effect of floret size on diversity of foraging bees was found, but associations of bee tongue length and floret size led to differences in the communities foraging on sunflowers with small, medium, or large florets. Though most wild bees preferred to forage on plants with more nectar and smaller florets (≈ easier access to nectar), Bombus spp. showed an opposite response to floret size, foraging preferentially on lines with large florets. Changes in Melissodes spp. foraging preference in the presence of Bombus spp. also suggest interactions between small- and large-bodied bees may be important.
Similar content being viewed by others

1 Introduction
Sunflowers, Helianthus spp., are native to North America and comprise more than 50 wild species (Rogers et al. 1982), most of which require cross-pollination due to genetic self-incompatibility (Heiser et al. 1969). Cultivated sunflower, Helianthus annuus L., is grown as a global oilseed crop that has been bred for improved self-fertility (i.e., reduced pollinator-dependence) for decades. However, the crop is still reliant on pollinators in at least two ways. First, to produce the hybrid cultivars sold by seed companies, pollen must be moved by bees from male-sterile to male-fertile inbred lines (DeGrandi-Hoffman and Chambers 2006, Greenleaf and Kremen 2006). Second, even within hybrid cultivars bred for self-fertility, wild and managed bees increase yields as much as 70% (Aslan and Yavuksuz 2010a, b; Carvalheiro et al. 2011; Mallinger et al. 2019; Mallinger and Prasifka 2017b). The observed increase in yield for self-fertile hybrids that are allowed open-pollination by insects may be related to plant stress due to high temperatures during bloom (DeGrandi-Hoffman and Chambers 2006).
Because of the ongoing importance of bees to sunflower production, research in sunflower-pollinator interactions has focused on determining what plant traits might explain variation in bee preference among cultivated sunflower inbreds and hybrids, with various studies indicating volatile compounds (Pham-Delégue et al. 1989), pollen availability (Tepidino and Parker 1982; Mallinger and Prasifka 2017a), and nectar sugar content or composition (Mallinger and Prasifka 2017a) as important traits. Most recently, wild bee visitation to sunflower inbred lines was strongly associated with ease of access to nectar, as indicated by the size (≈ depth) of florets (Mallinger and Prasifka 2017a; Portlas et al. 2018). Given that floret depth in sunflower is controlled by a small number of genes which do not appear to be linked to other key traits (Reinert et al. 2020), there is potential to breed for sunflowers with shorter florets as preferred by nectar-foraging bees.
Recent studies on sunflower pollination also have found that an assortment of native, wild bees, rather than honeybees, Apis mellifera L., account for most of the pollination services provided to sunflowers in the Midwestern USA (Mallinger and Prasifka 2017a, 2017b; Mallinger et al. 2019; Portlas et al. 2018). Though there is variation in tongue length for honey bee workers (Waddington and Herbst 1987), the broader native bee community shows far greater variation in tongue length which is expected to impact their foraging preferences (Thompson 2001, Rollings and Goulson 2019). In spite of the variation in body size and tongue length, native bees as a community still show greater visitation rates to sunflowers with shorter florets (Mallinger and Prasifka 2017a; Portlas et al. 2018). Though Mallinger and Prasifka (2017a, 2017b) and Mallinger et al. (2019) identified most bees to species level, they did not evaluate possible effects of floret size on pollinator community composition and diversity.
Research on sunflower pollination has emphasized the effects of specific plant traits on pollinator visitation rates or abundance but yields of pollinator-dependent crops sometimes depend on the diversity of bee visitors rather than their abundance (Hoehn et al. 2008). While previous work identified greater bee visitation rates to shorter florets (Mallinger and Prasifka 2017a; Portlas et al. 2018), whether or not floret length influenced overall bee diversity or community composition was not explored. Accordingly, this study examined whether sunflower lines with small, medium, or large florets were visited by different bee communities. Estimates of bee tongue lengths for the different species found visiting sunflowers were also used to evaluate if tongue lengths matched the floret sizes of their preferred sunflower lines.
2 Materials and methods
2.1 Field site
Inbred sunflower lines were planted on May 16, and June 14 in 2018 and May 21 and June 14 in 2019 at Casselton, ND, under standard cultivation practices. Each row was 6 m in length with 0.76 m between rows. Experimental plots were part of larger sunflower fields with dimensions of 52 × 128 m in 2018 and 49 × 122 m in 2019. Six lines were planted in 2018 and increased to 12 lines in 2019, including three wild annual sunflower accessions (entries held in the USDA’s National Plant Germplasm System). The single row plots in 2018 spanned 18 rows (in a single range) in the early planting and 6 rows in 3 ranges in the late planting (4 large, 2 medium, and 3 small varieties). In 2019, plots were 12 rows in two ranges (3 large, medium, small, and wild varieties) in both early and late plantings. Additional lines in 2019 were selected to balance sampling in floret size categories and maximize overlap of bloom across the lines. Each line was planted as a two-row pair and each pair considered a unit for sampling and analysis purposes. Floret depths were categorized as small (< 7.5 mm), medium (7.5–9 mm), or large (> 9 mm).
2.2 Floret collection
Floret samples were taken by cutting a wedge from the head representing roughly one-sixth of the total area. To standardize location of sampled florets, wedges were taken when 10–30% of florets had begun anthesis. Wedges were stored in the freezer (− 18 °C) until processed. Florets were removed from the wedge and scanned, and TIFF images were generated. Using ImageJ (Schneider et al. 2012), a macro (automated code block) was created that measured the length of each floret.
2.3 Bee collection and identification
To evaluate the community composition and diversity of bees foraging across sunflower lines, a standardized number of foragers were collected across lines. In the 2018 season, up to 30 bees were collected during bloom for each line, and in 2019, a total of 25 bees per line were collected over 5 concurrent days. Bees were collected after they had begun foraging on a sunflower head. Bees were collected individually in 50 ml vials, kept in a cooler on ice packs until they could be brought out of the field, and then stored in a freezer at − 18 °C until they were processed. Specimens were pinned and identified using DiscoverLife© keys (Ascher and Pickering 2017). Species were verified using a reference collection of bees from sunflowers in the Great Plains. Specimens in Lasioglossum and Diadasia were identified to genera and all other bees were identified to species. Lasioglossum and Diadasia represented 1–2% of the specimens collected, so it was determined genus level identification was sufficient. Specimens are currently stored at USDA ETSARC in Fargo, ND.
2.4 Walking bee counts
To quantify visitation rates across lines, at the onset of bloom in 2019, bee counts were conducted for each sunflower line by walking between the pair of rows per line and counting bees foraging on the heads. Each surveying event for a line was 1 min in duration and was repeated 4 times, approximately 1 h apart over a 4-h period. Wild sunflowers were not included in the walking counts due to differences in the number and size of heads per plant (making it challenging to compare visitation rates) and a bloom period that extended before and after the inbred lines.
2.5 Nectar sampling
Within each sunflower line, a subsample of n = 3 sunflower heads were bagged when the plants had 10% of florets in anthesis. Bags were secured to prevent bees and other insects from foraging on the florets. A 6.6-μl microcapillary tube was used to sample nectar from individual florets by inserting the tube until it reached the base of the floret. Florets sampled were 24 h post-anthesis, and 10 florets were sampled per head or until nectar volume approached the maximum capillary height of 72 mm. For each sample of ~ 10 florets, the length of the tube containing nectar was measured in millimeters, and nectar volume per floret in mm was determined by dividing nectar volume by the number of florets sampled (less than or equal to 10). The nectar within the tube was immediately dispensed onto a handheld refractometer and the percentage of sugar (Brix) in the nectar recorded.
2.6 Statistical analysis
All data were analyzed using RStudio 1.1.463 (RStudio Team 2018). Species diversity per sunflower line was estimated with SpadeR (Chao et al. 2015) using bootstrapping (n = 100). Hill numbers (effective number of species) (q = 0, q = 1, q = 2) were used to compare diversity between floret categories (short, medium, large) within each year × planting date. Sensitivity to species relative abundances is indicated by “q” where more weight is given to the most abundant species as “q” increases (Chao et al. 2014a). Hill numbers are standardized to allow for quantification of species and comparison across multiple systems (Chao et al. 2014b), and do not require abundances to be equal (Harvey et al. 1983). Diversity values were compared between floret categories with an ANOVA in RStudio. The similarity of bee communities visiting different floret size categories was assessed by conducting a PERMANOVA (package “vegan”) on the species abundances with planting date, floret category, and planting date × floret category as the factors.
Non-metric multidimensional scaling (NMDS) was conducted using package “vegan” and function metaMDS based on Bray–Curtis similarity and two dimensions. Comparisons of relative species abundance between planting dates were performed for each year with the addition of floret size as a factor. The significance of vectors was determined using permutations of 999 random permutations. Only significant variables were demonstrated (p < 0.05).
To examine the relationship between bee tongue length and floret length, tongue lengths were estimated for each specimen from morphometrics and models built by Cariveau et al. (2016) (package = ‘BeeIT). For each year × planting date combination, a Kruskal–Wallis one-way analysis of variance tested for differences in bee tongue length distributions across floret size categories. Dunn’s test (package = “dunn.test”) (Dinno 2017) was used to make pair-wise multiple comparisons post hoc (significance occurs at alpha/2 where alpha = 0.05). Calculations to obtain the mismatch between bee tongue length and floret size were taken by subtracting the floret size from the tongue length of each individual bee (Stang et al. 2009). When values are < 0, the floret is longer than the tongue and when > 0 the floret is shorter. Of the species collected, only four were present in sufficient numbers in both the early and late plantings to analyze differences. Average values were compared between early and late-collected Melissodes agilis Cresson, M. communis Cresson, M. trinodis Robertson, and Lasioglossum spp. using Welch two sample t-test. There were four species of Bombus that had enough specimens to determine average degree of mismatch: Bombus bimaculatus Cresson, B. griseocollis (De Deer), B. impatiens Cresson, and B. ternarius Say. Because the Bombus species were not present in the early planting, mismatch could only be calculated for the late planting.
Stepwise forward regression (package = “MASS”) (Ripley et al. 2019) was conducted to determine the relationship between bee visitation rates determined in timed walking counts and floral traits including floret length, nectar volume, and nectar concentration for the two different plantings (early and late) in 2019. This analysis was conducted using total visitation rates in early planted sunflower for all bees combined, and for Bombus spp. and non-Bombus spp. separately. Bees were separated into two groups in the late planting due to the majority of Bombus spp. only being present in the later planting.
3 Results
3.1 Bee diversity in relation to floret size
The number of species observed per sunflower line varied between 2 and 8, with an average of 5 species collected. In 2018, Melissodes agilis and M. trinodis were the most abundant bees in both the early and late planted sunflowers (Table I). A similar pattern was seen in 2019 (M. agilis and M. trinodis were the most abundant in early and in small and medium florets late), but Bombus griseocollis was most abundant in large florets of late planted sunflowers (Table II). No evidence for floret size affecting diversity of foraging bees was found, as species richness (q = 0) (early 2018: F(2,4) = 1.18, p = 0.40; late 2018: F(2,6) = 1.36, p = 0.33; early 2019: F(2,9) = 1.23, p = 0.37; late 2019: F(2,9) = 0.33, p = 0.73), Shannon diversity (q = 1) (early 2018: F(2,4) = 2.42, p = 0.21; late 2018: F(2,6) = 2.61, p = 0.15; early 2019: F(2,9) = 0.97, p = 0.42; late 2019: F(2,9) = 0.23, p = 0.79), and Simpson diversity (q = 2) (early 2018: F(2,4) = 2.07, p = 0.24; late 2018: F(2,6) = 1.97, p = 0.22; early 2019: F(2,9) = 0.62, p = 0.56; late 2019: F(2,9) = 0.15, p = 0.86) were not significantly different (p > 0.05) among floret sizes for any year × planting date combination (Figure 1).

Diversity curves constructed with interpolated (solid line) and extrapolated (dashed line) data. Bands indicate a 95% confidence interval. Curves were plotted for small, medium, and large florets for: early 2018 (A); late 2018 (B); early 2019 (C); late 2019 (D)
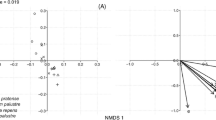
The NMDS illustrated that the composition of bee communities visiting sunflowers was not affected by floret size in 2018, but communities differed between early and late plantings (F(1,16) = 9.02, p = 0.001). For 2019, both date (F(1,23) = 6.09, p = 0.001) and floret size (F(2,23) = 2.48, p = 0.01) significantly influenced the composition of the sunflower bee communities (Figure 2).

Non-metric multi-dimensional scaling (NMDS) plots illustrating the distribution of species in early vs late plantings for 2018 (A) and 2019 (B). Ellipses represent 95% confidence intervals for each planting date’s centroid. Stress values indicate a fair to good fit
3.2 Bee tongue length in relation to floret size
Distributions of bee tongue lengths differed among the floret categories (small, medium, large) within both the early 2018 (χ2 = 15.35, df = 2, p < 0.01) and late 2018 (χ2 = 41.80, df = 2, p < 0.01) planting dates. The same general pattern was found in both early (χ2 = 28.78, df = 2, p < 0.01) and late (χ2 = 41.49, df = 2, p < 0.01) planting dates in 2019. Significant pairwise differences always occurred between small and large florets (p < 0.05), but differences in tongue length distributions between medium florets and the other size categories were inconsistent (Figure 3).

Distribution of bee tongue lengths by floret category: early 2018 (A), late 2018 (B), early 2019 (C), late 2019 (D). Within the year and planting date, significant differences between distributions of bee tongue lengths were observed between the three floret categories (p ≤ 0.05) with the exception of Medium-Large in early 2018 (p = 0.367) and Small-Medium in late 2018 (p = 0.398)
Comparisons of the average bee tongue length to the depth of the floret where bee species were collected showed significant differences between the early and late planting. On average, M. agilis (t = − 9.39, df = 195, p < 0.05) and M. trinodis (t = − 3.95, df = 143, p < 0.05) foraged on lines with florets that were shallower relative to tongue length in the late planting (Figure 4). Lasioglossum sp. foraged only on sunflowers with florets deeper than the tongue lengths (t = − 2.7, df = 4, p < 0.07). Bombus spp. were present exclusively in the late planting, and large positive mismatch values show all four species foraged on lines with florets that were shallow relative to tongue length (B. bimaculatus 8.42 mm, B. griseocollis 8.70 mm, B. impatiens 5.10 mm, and B. ternarius 5.36 mm).

Average mismatch for bees collected in 2019. Species marked with an asterisk indicate significant difference between mismatch of the early and late planting (p < 0.05). Bombus sp. consistently foraged on sunflower florets whose depth was less than the tongue length
3.3 Bee visitation and floral traits
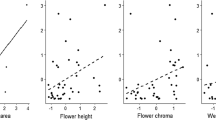
Bee responses to floral traits differed between planting dates in 2019. Forward stepwise regression showed bee visitation in early-planted sunflowers was significantly impacted by floret size, nectar volume, and sugar content (F(3,5) = 4.88, p = 0.05). In late-planted sunflower, the model excluding Bombus spp. only included nectar volume (F(3,5) = 6.70, p = 0.04) as a predictor. The separate model for Bombus spp. included nectar volume and floret size as significant factors (F(3,5) = 19.63, p < 0.01), with bees foraging preferentially on lines with large florets.
4 Discussion
Sunflowers in the northern Great Plains are typically planted between early May and mid-June, with decisions based on a variety of factors such as climate, convenience, and pest avoidance. Planting early means growers may harvest prior to inclement weather in the fall, and can also reduce damage from the red sunflower seed weevil (Smicronyx fulvus LeConte), the most damaging insect pest in this region (Oseto et al. 1987). Conversely, late plantings can help avoid damage by several other insect pests (Oseto et al. 1989; Prasifka et al. 2016). The short active periods for various wild bee species in summer mean changes in the bee community are likely between early- and late-planted sunflowers.
Understanding pollinator responses to crop traits in pollinator-dependent plants is important for maximizing and maintaining consistency of yields (Aizen et al. 2008; Hoehn et al. 2008). It is well-documented that bee foraging is influenced by the size of flowers (Harder 1983; Eckhart 1991; Conner and Rush 1996; Stang et al. 2009) and the quality or quantity of nectar (Harder 1983; Klumpers et al. 2019). Prior research in sunflowers has shown bees respond negatively to increasing floret depth and positively to increasing nectar volume (Mallinger and Prasifka 2017a, b, Mallinger et al. 2019, Portlas et al. 2018, Prasifka et al. 2018). Our findings in this study are similar to prior research and improve our understanding of how floret size and planting date influence the communities of bees pollinating sunflowers, and how specific pollinator responses to plant traits may differ. Further examination of bee species present in areas where sunflowers are cultivated (i.e., South Dakota, California, Texas) could help inform decisions on planting dates: if small- and medium-bodied bees (sunflower specialists) are considered more important, earlier planting dates may enhance pollination, but if large-bodied generalists (Bombus spp.) are needed, later planting would be beneficial.
Over 4 year × planting date combinations, collections of bees from sunflowers show two obvious patterns. First, M. agilis and M. trinodis were almost always the most abundant species, which is unsurprising given their status as sunflowers oligoleges (Parker et al. 1981; Ascher and Pickering 2017) and recent collections of sunflower pollinators in the Northern Great Plains (Mallinger and Prasifka 2017a, 2017b). Second, the numbers of Bombus spp., particularly B. griseocollis, increased greatly between bloom of early- and late-planted sunflowers. Bombus griseocollis was the most abundant species in lines with large florets for the late planting in 2019, perhaps because its long tongue is more suited to reach nectar at the bottom of large florets. The increase in Bombus spp. late in the season is a substantial change to the community of foraging bees in sunflowers, and studies focusing solely on early-planted sunflowers risk missing this genus and any effects these species have on pollination.
When pollinator diversity was assessed using various indices, floret size did not significantly affect the diversity of bees visiting sunflowers, with approximately ~ 5 bee species actively foraging in each floret size category. Despite an absence of floret size effects on diversity, the composition of foraging bee communities varied between planting dates and, in 2019, bee community composition differed between the floret sizes as well. The relative stability in species diversity across floret lengths and planting dates is beneficial given evidence that increased diversity is associated with greater crop yields (Blüthgen and Klein 2011; Garibaldi et al. 2014; Eeraerts et al. 2020).
Data on the distributions of bee tongue lengths provide another way to examine pollinator responses to floret depth. One generalization is that the ranges of tongue lengths were broad across floret size categories. Stated another way, bees foraged on a greater range of floret depths than would be expected based on tongue length and floret depth alone. Some of this variation can be attributed to aspects of floret morphology not accounted for in this study such as floral opening width and diameter (Eckhart 1991) relative to width of the head of the bee. It was also uncommon to find bees with the longest tongues (Bombus spp.) foraging in lines with small or medium florets; this may be a result of increased handling time that makes foraging on smaller florets less efficient for larger bees (Inouye 1980; Harder 1983; Klumpers et al. 2019). Even with a broad range of tongue lengths collected across floret size categories, there were still significant differences in distributions, with bees matching their body size to floret size (i.e., more short-tongued bees on lines with small florets). The trend towards foraging bees’ matching tongue lengths to floret size is another way to visualize and understand floret size effects on sunflower bee communities. Lastly, a significant effect of time can be seen by examining the average mismatch of tongue length and floret size. Specifically, while M. agilis and M. trinodis were always estimated to be foraging on florets whose depth was less than their tongue lengths, there was a significant increase in this mismatch for both species between the early- and late-planted sunflowers. Though not conclusive, the movement of M. agilis and M. trinodis towards smaller florets coinciding with the increase in Bombus spp. foragers is consistent with the idea that resource partitioning is occurring in the native bee community foraging on sunflowers.
Stepwise models of bee visitation to individual lines in 2019 provided another clarification of prior results in sunflowers. As seen by Mallinger and Prasifka (2017a), increasing nectar volume consistently elicited a positive response from bees across the season, and in early-planted sunflowers bee visitation decreased with increasing floret depth as observed by Portlas et al. (2018). However, Bombus spp. (only abundant in late-planted sunflowers) had the opposite response, with visitation increasing to lines with larger florets. This has been shown in other studies of bumble bees (Eckhart 1991; Conner and Rush 1996; Stout 2000) and, as noted above, may be a function of handling-time and related energetic costs.
Collectively, these results provide clarification of previous observations and recommend changes or future areas of study. To obtain a more complete understanding of bee pollination, future studies should attempt to include late-planted sunflowers, for which insect-plant interactions may differ due to the increased numbers of Bombus spp. While specific plant traits (floret size and other nectar-related traits) can explain most variation in bee foraging in sunflowers, additional factors are likely important in some contexts. In particular, interspecific interaction between small- and large-bodied bees is one likely source of variation in pollinator choice and should be further studied to understand its impact on insect behavior and crop pollination.
Data Availability
Data used in this manuscript can be made reasonably available upon request.
Code availability
Not applicable.
References
Aizen A., L. A. Garibaldi, S. A. Cunningham, and A. M. Klein. (2008) Long-term global trends in crop yield and production reveal no current pollination shortage but increasing pollinator dependency. Curr. Biol. 18 (20), 1572–1575
Alexis Dinno. (2017) Dunn’s test of multiple comparisons using rank sums
Ascher, J. S. and J. Pickering. (2017) DiscoverLife bee species guide and world checklist (Hymenoptera: Apoidea)
Aslan, M. M. and C. Yavuksuz. (2010) Effect of honeybee (Apis mellifera) and bumblebee (Bombus terrestris L.) pollinators on yield and yield factors in sunflower (Helianthus annuus L.) production areas. J. Anim. Vet. Adv. 9 (2), 332–335
Blüthgen, N., and A.-M. Klein. (2011) Functional complementarity and specialisation: The role of biodiversity in plant–pollinator interactions. Basic Appl. Ecol. 12, 282–291
Cariveau, D. P., G. K. Nayak, I. Bartomeus, J. Zientek, J. S. Ascher, J. Gibbs, and R. Winfree. (2016). The allometry of bee proboscis length and its uses in ecology. PLOS ONE. 13(11): e0207900. https://doi.org/10.1371/journal.pone.0207900
Carvalheiro, L. G., R. Veldtman, A. G. Shenkute, G. B. Tesfay, C. W. W. Pirk, J. S. Donaldson, and S. W. Nicolson. (2011) Natural and within-farmland biodiversity enhances crop productivity. Ecol. Lett. 14 (3), 251–259
Chao, A., C. H. Chiu, and L. Jost. (2014a) Unifying species diversity, phylogenetic diversity, functional diversity, and related similarity and differentiation measures through Hill numbers. Annu. Rev. Ecol. Evol. S. 45, 297-324
Chao, A., K. H. Ma, T. C. Hsieh, and C. H. Chiu. (2015) Introduction to online program SpadeR (Species-richness Prediction and Diversity Estimation in R)
Chao, A., N. J. Gotelli, T. C. Hsieh, E. L. Sander, K. H. Ma, R. K. Colwell, A. M. Ellison. (2014b) Rarefaction and extrapolation with Hill numbers: a framework for sampling and estimation in species diversity studies. Ecol. Monogr. 84, 45–67.
Rogers, C. E., T. E. Thompson, and G. J. Seiler. (1982) Sunflower species of the United States. National Sunflower Association, Fargo, ND 58102 U.S.A.
Conner, J. K., and S. Rush. (1996) Effects of flower size and number on pollinator visitation to wild radish, Raphanus raphanistrum. Oecologia. 105 (4), 509–516
Degrandi-Hoffman, G. and M. Chambers. (2006) Effects of honeybee (Hymenoptera: Apidae) foraging on seed set in self-fertile sunflowers (Helianthus annuus L.). Environ. Entomol. 35 (4), 1103–1108
Eeraerts, M., G. Smagghe, and I. Meeus. (2020) Bumble bee abundance and richness improves honeybee pollination behaviour in sweet cherry. Basic Appl. Ecol. 43, 27–33
Eckhart, V. M. (1991) The effects of floral display on pollinator visitation vary among populations of Phacelia linearis (Hydrophyllaceae). Evol. Ecol. 5, 370–384
Garibaldi, L. A., L. G. Carvalheiro, S. D. Leonhardt, M. A. Aizen, B. R. Blaauw, et al. (2014) From research to action: enhancing crop yield through wild pollinators. Front. Ecol. Environ. 12 (8), 439–447
Greenleaf, S. S., and C. Kremen. (2006) Wild bees enhance honeybees’ pollination of hybrid sunflower. P. Natl. Acad. Sci. 103 (37), 13890–13895
Harder, L. D. (1983) Flower handling efficiency of bumble bees: Morphological aspects of probing time. Oecologia. 57, 274-280
Harvey, P. H., R. K. Colwell, J. W. Silvertown, and R. M. May. (1983) Null models in ecology. Annu. Rev. Ecol. Syst. 14, 189-211
Heiser, C. B., D. M. Smith, S. B. Clevenger, and W. C. Martin. (1969) The North American Sunflowers (Helianthus). Memoirs of the Torrey Botanical Club. 22, 1–218
Hoehn, P., T. Tscharntke, J. M. Tylianakis, I. Steffan-Dewenter. (2008) Functional group diversity of bee pollinators increases crop yield. P. Roy. Soc. B. 275 (1648): 2283-2291
Inouye, D. W. 1980. The effect of proboscis and corolla tube lengths on patterns and rates of flower visitation by bumblebees. Oecologia. 45, 197–201
Klumpers, S. G. T., M. Stang, and P. G. L. Klinkhamer. (2019) Foraging efficiency and size matching in a plant-pollinator community: the importance of sugar content and tongue length. Ecol. Lett. 22 (3), 469-479
Mallinger, R. E., J. Bradshaw, A. J. Varenhorst, and J. R. Prasifka. (2019) Native solitary bees provide economically significant pollination services to confection sunflowers (Helianthus annuus L.) (Asterales: Asteraceae) grown across the Northern Great Plains. J. Econ. Entomol. 112 (1), 40–48
Mallinger, R. E., and J. R. Prasifka. (2017a) Bee visitation rates to cultivated sunflowers increase with the amount and accessibility of nectar sugars. J. of Appl. Entomol. 141 (7), 561–573
Mallinger, R., and J. Prasifka. (2017b) Benefits of insect pollination to confection sunflowers differ across plant genotypes. Crop Sci. 57 (6), 3264
Oseto, C. Y., Busacca, J. D., Charlet, L. D. (1987) Relationship of sunflower planting dates to damage by Smicronyx fulvus (Coleoptera: Curculionidae) in North Dakota. J.Econ. Entomol. 80 (1), 190–192.
Oseto, C. Y., Charlet, L. D., Busacca, J. D. (1989) Effects of planting date on damage caused by the banded sunflower moth (Lepidoptera: Cochylidae) in the northern Great Plains. J. Econ. Entomol. 82 (3), 910–912.
Parker, F. D., V. J. Tepedino, and G. E. Bohart. (1981) Notes on the biology of a common sunflower bee, Melissodes (Eumelissodes) agilis Cresson. J. of the New York Entomol. Soc. 89 (1), 43–52
Pham-Delégue, M. H., O. Bailez, M. M. Blight, C. Masson, A. L. Picard-Nizou, and L. J. Wadhams. (1989) Sunflower volatiles involved in honeybee discrimination among genotypes and flowering stages. J. Chem. Ecol. 15 (1), 329–343
Portlas, Z. M., J. R. Tetlie, D. Prischmann-Voldseth, B. S. Hulke, and J. R. Prasifka. (2018) Variation in floret size explains differences in wild bee visitation to cultivated sunflowers. Plant Genet. Resour-C. 16 (6), 498–503
Prasifka et al. 2018: Prasifka, J. R., R. E. Mallinger, Z. M. Portlas, B. S. Hulke, K. K. Fugate, T. Paradis, M. E. Hampton, and C. J. Carter. (2018). Using nectar-related traits to enhance crop-pollinator interactions. Frontiers in Plant Science 9. https://doi.org/10.3389/fpls.2018.00812
Prasifka, J.R., L.F. Marek, D.K. Lee, S. Thapa, V. Hahn, and J.D. Bradshaw. (2016) Effects from early planting of late-maturing sunflowers on damage from primary insect pests in the United States. Helia 39 (64), 45-56.
Reinert, S., Q. Gao, B. Ferguson, Z. M. Portlas, Jarrad. R. Prasifka, and B. S. Hulke. (2020) Seed and floret size parameters of sunflower are determined by partially overlapping sets of quantitative trait loci with epistatic interactions. Mol. Genet. Genom. 295 (1), 143–154
Ripley, B., B. Venables, D. M. Bates, K. Hornik, A. Gebhardt, and D. Firth. (2019) Support functions and datasets for venables and Ripley’s MASS
Rollings and Goulson 2019: Rollings, R. and D. Goulson. (2019). Quantifying the attractiveness of garden flowers for pollinators. Journal of Insect Conservation 23: 803-817
RStudio Team. (2018) RStudio: Integrated Development for R. RStudio, Inc., Boston, MA
Schneider, C. A., W. S. Rasband, and K. W. Eliceiri. (2012) NIH Image to ImageJ: 25 years of image analysis. Nat. Methods. 9, 671–675
Stang, M., P. G. L. Klinkhamer, N. M. Waser, I. Stang, E. Meijden. (2009) Size-specific interaction patterns and size matching in a plant-pollinator interaction web. Ann. Bot. 103 (9), 1459–1469
Stout, J. C. (2000) Does size matter? Bumblebee behaviour and the pollination of Cytisus scoparius L. (Fabaceae). Apidologie. 31 (1), 129–139
Thompson, J. D. (2001). How do visitation patterns vary among pollinators in relation to floral display and floral design in a generalist pollination system? Oecologia 126: 386-395
Tepidino, V. J., and F. D. Parker. (1982) Interspecific differences in the relative importance of pollen and nectar to bee species foraging on sunflowers. Environ. Entomol. 11 (1), 246–250
Waddington, K. D., and L. H. Herbst. (1987) Body size and the functional length of the proboscis of honeybees. Fla. Entomol. 70 (1), 124–128
Acknowledgements
We thank Lisa Brown (USDA), Ben Glogoza, Robert Brenner, and Shawna Pantzke (NDSU) for their help collecting insect specimens and plant material from the field. Additional thanks to Brian Otteson (NDSU) at the NDSU Agronomy Seed Farm for site selection and field preparation, as well as Brady Keohler and Mike Grove (USDA) for help with planting. Funding through USDA-ARS project 3060-21000-043-00-D, “Genetic Enhancement of Sunflower Yield and Tolerance to Biotic Stress.”
Funding
Research funding was provided by the USDA-ARS project 3060–21000-043–00-D, “Genetic Enhancement of Sunflower Yield and Tolerance to Biotic Stress.”
Author information
Authors and Affiliations
Contributions
Beth Ferguson and Jarrad Prasifka contributed to the study design and conception. Data collection and analysis were performed by Beth Ferguson. Bee identifications were confirmed by Rachel Mallinger. The manuscript was written by Beth Ferguson, and all authors commented on prior versions of the manuscript. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval
This research was conducted using an unregulated invertebrate species; therefore, approval from a research ethics committee was not required.
Consent to participate
Not applicable.
Consent for publication
Not applicable.
Conflict of interest
The authors declare no competing interests.
Additional information
Manuscript editor: Mathieu Lihoreau
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
About this article

Cite this article
Ferguson, B., Mallinger, R.E. & Prasifka, J.R. Bee community composition, but not diversity, is influenced by floret size in cultivated sunflowers. Apidologie 52, 1210–1222 (2021). https://doi.org/10.1007/s13592-021-00897-z
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s13592-021-00897-z