
Abstract
It has been proposed in the past that adaptation to rotated visual feedback is based on directionally tuned modules. Here, we investigate whether adaptation depends on the number of modules that are concurrently activated. To disambiguate the number of modules and their spatial overlap, we decided to vary the number of target directions and their spacing independently. In light of recent work on the existence of fast and slow adaptive processes, we analyzed the role of target number and spacing separately for the first eight movements under rotated visual feedback and for later movements. We found that during the first eight movements, adaptation progressed three times faster when targets were spaced across a 42° rather than across a 360° range, irrespective of their number. During the subsequent movements, adaptation progressed 1.66 times faster with two than with eight targets, irrespective of their spacing. This differential dependence of early and late adaptation on target metrics confirms the existence of adaptive processes with different time scales and suggests that those processes differ not only by their clocking speed, but also by their functional properties. Specifically, the speed of fast processes seems to be constrained by the directional tuning width of adaptive modules, and the speed of slow processes by the number of presentations per target direction.



Similar content being viewed by others
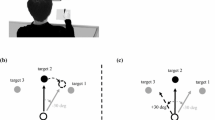


Notes
Randomization was performed with the in C++ statement “rand” after seeding with “srand”.
Movement onset was defined by a velocity threshold of 30 mm/s.
We decided for eight movements to represent fast adaptive processes since all target directions were presented equally often within eight movements (see "Procedures"). This is slightly less than the ten or twelve movements selected in earlier studies on adaptation to mirror reversals (Stratton et al. 2007) and force fields (Smith et al. 2006), respectively. Since the first eight adaptation movements in the present study took about 30 s, our choice is comparable to the value determined for fast adaptation to visual rotations in psychophysical experiments (Inoue et al. 2000). We are therefore confident that our analysis didn’t exceed the temporal limits of fast processes.
References
Abeele S, Bock O (2001) Mechanisms for sensorimotor adaptation to rotated visual input. Exp Brain Res 139:248–253
Andersen RA, Bracewell RM, Barash S, Gnadt JW, Fogassi L (1990) Eye position effects on visual, memory, and saccade-related activity in areas LIP and 7a of macaque. J Neurosci 10:1176–1196
Bock O, Abeele S, Eversheim U (2003) Human adaptation to rotated vision: interplay of a continuous and a discrete process. Exp Brain Res 152:528–532
Georgopoulos AP, Kettner RE, Schwartz AB (1988) Primate motor cortex and free arm movements to visual targets in three-dimensional space. II. Coding of the direction of movement by a neuronal population. J Neurosci 8:2928–2937
Ghahramani Z, Wolpert D (1997) Modular decomposition in visuomotor learning. Nature 386:392–395
Imamizu H, Uno Y, Kawato M (1995) Internal representations of the motor apparatus: implications from generalization n visuomotor learning. J Exp Psychol Hum Percept Perform 21:1174–1198
Inoue K, Kawashima R, Satoh K, Kinomura S, Sugiura M, Goto R, Ito M, Fukuda H (2000) A PET study of visuomotor learning under optical rotation. NeuroImage 11:505–516
Kalaska JF, Caminiti R, Georgopoulos AP (1983) Cortical mechanisms related to the direction of two-dimensional arm movements: relations in parietal area 5 and comparison with motor cortex. Exp Brain Res 51:247–260
Krakauer J, Pine Z, Ghilardi M-F, Ghez C (2000) Learning of visuomotor transformations for vectorial planning of reaching trajectories. J Neurosci 20:8916–8924
Lee J, Schweighofer N (2009) Dual adaptation supports a parallel architecture of motor memory. J Neurosci 29:10396–10404
Ottes F, Van Gibergen J, Eggermont J (1986) Visuomotor fields of the superior colliculus: a quantitative model. Vision Res 26:857–873
Roby-Brami A, Burnod Y (1995) Learning a new visuomotor transformation: error correction and generalization. Cogn Brain Res 2:229–242
Smith M, Ghazizadeh A, Shadmehr R (2006) Interacting adaptive processes with different timescales underlie short-term motor learning. PLoS Biol 4:1035–1043
Snoddy G (1926) Learning and stability. J Appl Psychol 10:1–36
Stratton S, Liu Y-T, Hong S, Mayer-Kress G, Newell K (2007) Snoddy (1926) revisited: time scales of motor learning. J Motor Behav 39:503–515
Tanaka H, Sejnowski T, Krakauer J (2009) Adaptation to visuomotor rotation through interaction between posterior parietal and motor cortical areas. J Neurophysiol 102:2921–2932
Wang J, Sainburg R (2005) Adaptation to visuomotor rotations remaps movement vectors, not final positions. J Neurosci 25:4024–4030
Woolley D, Tresilian J, Carson R, Rick S (2007) Dual adaptation to two opposing visuomotor rotations when each is associated with different regions of workspace. Exp Brain Res 179:155–165
Acknowledgments
This work was supported by the German Space Agency DLR on behalf of the Bundesministeriu für Wirtschaft und Technologie (Grant 50WB0625). Thanks are due to Dipl.-Ing. (FH) L. Geisen for software development and to V. Rohde, K. Friederichs, and M. Hinrichs for their help with data collection and analysis.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article
Cite this article
Bock, O., Schmitz, G. Adaptation to rotated visual feedback depends on the number and spread of target directions. Exp Brain Res 209, 409–413 (2011). https://doi.org/10.1007/s00221-011-2564-8
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00221-011-2564-8