
Abstract
Many everyday activities require coordination and monitoring of multiple deadlines. One way to handle these temporal demands might be to represent future goals and deadlines as a pattern of spatial relations. We examined the hypothesis that spatial ability, in addition to executive functioning, contributes to individual differences in multitasking. In two studies, participants completed a multitasking session in which they monitored four digital clocks running at different rates. In Study 1, we found that individual differences in spatial ability and executive functions were independent predictors of multiple-task performance. In Study 2, we found that individual differences in specific spatial abilities were selectively related to multiple-task performance, as only coordinate spatial processing, but not categorical, predicted multitasking, even beyond executive functioning and numeracy. In both studies, males outperformed females in spatial ability and multitasking and in Study 2 these sex differences generalized to a simulation of everyday multitasking. Menstrual changes moderated the effects on multitasking, in that sex differences in coordinate spatial processing and multitasking were observed between males and females in the luteal phase of the menstrual cycle, but not between males and females at menses. Overall, these findings suggest that multiple-task performance reflects independent contributions of spatial ability and executive functioning. Furthermore, our results support the distinction of categorical versus coordinate spatial processing, and suggest that these two basic relational processes are selectively affected by female sex hormones and differentially effective in transforming and handling temporal patterns as spatial relations in the context of multitasking.
Similar content being viewed by others

Multitasking implies dealing with multiple goal-directed tasks and is required in order to successfully navigate through numerous everyday activities. A core feature of multitasking is the requirement for temporal integration and monitoring of overlapping action sequences within limited time frames. How people allocate limited cognitive resources to multiple concurrent tasks is a topic of considerable theoretical and practical interest (e.g., Burgess, Veitch, de Lacy Costello & Shallice, 2000; Fuster, 1993; Logie, Trawley & Law, 2011; Mäntylä, 2013; Meyer & Kieras, 1997; Navon & Gopher, 1979; Norman & Bobrow, 1975; Salvucci & Taatgen, 2008; Todorov, Del Missier & Mäntylä, 2014; Wickens, 2002).
Multitasking is a complex construct encompassing many different conditions with regard to time span, number of tasks to be completed and task difficulty, among others. However, a core aspect is represented by the ability to monitor concurrently multiple deadlines and to switch timely between tasks to meet the different deadlines (see also Mäntylä, 2013). This represents a fundamental aspect of multitasking, common to diverse situations observed in different contexts. As a consequence, the ability to represent and handle multiple concurrent deadlines seems to be a critical aspect of multitasking.
We have recently suggested (Mäntylä, 2013; Mäntylä & Todorov, 2013; Todorov et al., 2014) that multiple-task performance reflects individual differences in two fundamental aspects of cognitive functioning, namely, executive control and spatial ability. In this framework, and consistent with earlier work, variability in prefrontally mediated executive functions is considered as the primary source of individual differences in multitasking (Logie et al., 2011; Meyer & Kieras, 1997; Salvucci & Taatgen, 2008; Shallice & Burgess, 1991; Watson & Strayer, 2010).
We also proposed a spatiotemporal hypothesis positing that multitasking performance reflects individual differences in spatial processing. The spatiotemporal hypothesis reflects the idea that, under high demands on temporal coordination (e.g., multiple tasks with narrow deadlines), individuals with efficient spatial abilities should be better multitaskers than less spatially-skilled individuals. Most goal-directed tasks, including multiple-task performance, are temporal in that scheduling, monitoring and task interleaving take place on a (relative or absolute) time scale, and that coordinating multiple goals and deadlines requires a high degree of cognitive control. We suggested that one way to handle these executive control demands is to represent the temporal pattern of deadlines and task goals in spatial terms (e.g., Bonato, Zorzi & Umiltà, 2012; Casasanto & Boroditsky, 2008; Dehaene & Brannon, 2011).
As a support for this line of reasoning, Mäntylä (2013; see also Todorov et al., 2014) reported a study in which individuals with varying spatial abilities completed a multitasking session with four monitoring tasks and separate tasks of executive functioning and spatial ability. Individual differences in executive functioning (working memory updating) and spatial ability (mental rotation) were shown to be independent predictors of multiple-task monitoring, but only spatial ability was related to sex differences in multitasking. Inconsistent with popular beliefs and media claims (Fisher, 1999; Pease & Pease, 2003), the findings of the study showed that males exceeded females in multitasking performance. Furthermore, menstrual fluctuation moderated these effects, in that significant sex differences in multitasking performance (and spatial ability) were observed between males and females in the luteal, but not in the menstrual, phase of the cycle. Additionally, sex differences in multitasking performance were fully mediated by spatial ability. These findings suggest that multitasking involves spatiotemporal task coordination and that sex differences in multiple-task performance reflect differences in spatial ability.
In the present study, we examined the generality of the spatiotemporal hypothesis by testing its predictions in two studies. In both studies, multitasking performance was assessed with four identical and simple component tasks, requiring a high degree of coordination among the tasks. In this counter task, participants have to monitor four digital “clocks” (counters) that are identical, in that they display forward-running digits, and instructions are to press the spacebar whenever one of the counters shows a target reading, which was defined by a simple rule (see also Mäntylä, 2013). It is worth pointing out that earlier work involving a single monitoring task of the same kind did not reveal sex differences in response accuracy or in monitoring frequency (Mäntylä, Carelli, & Forman, 2007; Mäntylä, Del Missier, & Nilsson, 2009; see also e.g., Ceci & Bronfenbrenner, 1985).
This task represents a well-controlled paradigm, which captures essential aspects of multitasking (i.e., simultaneously monitoring multiple deadlines and switching timely between tasks), while eliminating or at least attenuating several confounds that usually affect more realistic multitasking situations, in which component tasks have domain-specific and task-specific requirements, and thus possibly require a combination of diverse skills that do not pertain to multitasking per se.
In Study 1, we examined the generality of our earlier findings, which were based on selected samples of Swedish undergraduates, by involving a more heterogeneous sample of Italian participants. Given the large ongoing discussion about replication in psychology (Pashler & Wagenmakers, 2012; see the whole issue of the journal dedicated to replication in psychological science), we aimed to replicate the study of Mäntylä’s (2013) in a different country. We also examined sex differences in multitasking by involving a more representative sample of female participants in that they were recruited independent of their menstrual and hormonal status.
In Study 2, we further examined the spatiotemporal hypothesis of multitasking by focusing on specific spatial processes underlying multitasking performance. Mäntylä (2013) used the Mental Rotations Test (MRT; Vandenberg & Kuse, 1978, Peters et al., 1995) to find that individual differences in spatial ability are related to multiple-task performance. While MRT is a widely used test of spatial ability, differences in mental-rotation scores do not provide a precise identification of the nature of spatial processes supporting multitasking. Indeed, even if MRT can be used as a proxy to show that spatial ability contributes to multitasking, finding a positive association between MRT and counter performance does not tell much about the specific spatial processes underlying individual and sex-related differences in multitasking.
With the aim of investigating this latter issue, we adopted the view that processing of spatial relations reflects two distinct aspects, often referred to as coordinate and categorical spatial processing (or absolute vs. relative processing; e.g., Kosslyn, 1987; Kosslyn et al., 1989; see also see Jager & Postma, 2003; Laeng, Chabris & Kosslyn, 2003; for overviews). Building on this distinction, we hypothesized that that individual differences in spatial ability reflect selective effects of coordinate (vs. categorical) spatial processing, and that these selective effects contribute to individual and sex-related differences in multitasking (see the introduction of Study 2 for a more detailed articulation of these arguments).
The second aim of Study 2 was to examine the external validity of our findings. Specifically, because the primary focus of this study was on individual and sex-related differences in the basic mechanisms of higher-order task coordination, it was necessary to use (gender-fair) test conditions in a well-controlled multitasking paradigm (i.e., the counter task), in which the role of domain-specific skills and experiences were minimized. As these positive aspects related to task control increase internal validity, they might actually reduce external validity. With this in mind we also related counter-task performance to a more realistic simulation of everyday multitasking (see Study 2 for details). We expected the two tests of multitasking to be positively correlated and to show similar sex differences. We also expected that individual differences in spatial ability should contribute to multitasking performance in both tasks.
Study 1
Method
Participants
The sample consisted of 102 young adults (42 males and 60 females) with the mean age of 22.57 years (SD = 3.37), ranging between 18 and 38 years of age. Sample size was determined on the basis of our previous studies with similar tasks (Mäntylä, 2013; Todorov et al., 2014). A large majority of the participants were University of Trieste undergraduates, and the remaining participants were young adults with a similar level of education and socio-economic status as the student sample.
Tasks and procedure
Spatial ability was assessed with the MRT (Vandenberg & Kuse, 1978; Peters et al., 1995). On each trial, participants are presented a target figure, placed in the left part of the sheet, and four possible alternatives of response, positioned on the right. The task is to identify the two figures, among the four presented, which represent the rotated target figure. Participants are asked to solve correctly the greatest number of problems in the given time limit. We used sections A and D of the MRT and participants had 3 min to complete each section of 12 problems. A short break took place between the two sessions. A response was scored as correct only if both stimulus figures were correctly identified. The final score was the sum of correct responses.
Executive functioning was assessed with a letter memory task, which measures the ability to update working memory contents, which together with the ability to shift between different mental sets and the inhibition of dominant or prepotent responses, is considered as a core component of executive functioning (Miyake et al., 2000). On each of the 14 trials, a sequence of letters, varying randomly in length between 5 to 12 items, was presented at the rate of 2 s/item on the computer screen (see also Del Missier, Mäntylä, & Bruine de Bruin, 2012). Participants were instructed to report the last three letters of each sequence. Two training series were also included to ensure that participants understood the task instructions, and a short break was allowed after the first half of the test sequences. The final score was the number of correctly reported triplets.
Multitasking was assessed with the counter task in both studies. In this time-based monitoring task, four digital clocks, or counters, were occluded by colored rectanglesFootnote 1 on the computer screen (see also Mäntylä, 2013). Participants could monitor each counter by pressing a specific key, whereupon the corresponding counter appeared for 2 s. To prevent the four tasks from being handled as a unitary task, the counters ran at different rates (4.2 s, 3.7 s, 2.7 s, and 2 s per item, respectively). Participants pressed spacebar whenever one of the counters displayed a target reading defined by a simple rule. Participants were instructed to press the spacebar when the last digit of the Green Counter (running at 4.2 s/item) was 7, when the last two digits of the Blue Counter were a multiple of 11, when the last two digits of the Red Counter were a multiple of 20, and when the last two digits of the Yellow Counter (running at 2 s/item), were a multiple of 25. Participants could check the reading of each counter whenever they wanted by pressing a designated key on the keyboard. The task took 20 min to complete and response accuracy (max = 107/20 min) and monitoring frequency were the dependent measures.
Participants were tested individually, during a single laboratory session. Informed consent was verbally obtained before participation, and the study was completed according to the APA ethical guidelines. Participants first completed a socio-demographic background questionnaire and the MRT, followed by the computerized letter-memory and counter tasks, administered in that order.
Results and discussion
Multitasking performance was based on a combined score of the four counter tasks (see also Mäntylä, 2013), with response accuracy and monitoring frequency as dependent measures. As the latter measure did not show any systematic effects (or trade offs, see Mäntylä et al., 2009), accuracy was the primary measure of counter task performance. A response was considered as correct if the spacebar was pressed within one digit of the target (e.g., the digits of 19, 20, and 21 would be considered correct responses if the target was 20). Due to technical problems, counter task data were not obtained for two participants (N = 99 for this task).
Table 1 summarizes the correlation data for the measures of spatial ability and executive functioning. Counter accuracy correlated significantly with MRT, providing additional support for the hypothesis that multitasking performance is related to individual differences in spatial ability. The correlation data also showed that letter memory scores were positively related to multitasking performance but not to MRT, possibly due to the verbal nature of the updating task (cf., Salway & Logie, 1995; Todorov et al., 2014). A regression analysis showed that both MRT (Beta = .26, p < .01) and letter memory (Beta = .24, p < .01), were significant predictors of counter accuracy.
The second main finding of Study 1 was that sex differences were observed in both multitasking, F(1, 99) = 18.29, η 2 = .16, p < .01, and mental rotation, F(1, 102) = 14.27, η 2 = .12, p < .01. As shown in Table 2, the sex difference in favor of males was apparent both in counter accuracy and MRT. By contrast, and consistent with past studies, executive functioning, as measured by the letter-memory task, did not show sex differences (F< 1).
The findings of Study 1 (and those of Mäntylä, 2013) suggest that individuals who are good at mental rotation are also better multitaskers than individuals with less efficient spatial abilities. These effects were consistent and replicated in different settings and samples of participants. However, MRT scores do not allow an accurate identification of the nature of spatial processes supporting individual and sex-related differences in multitasking performance. As MRT reflects multiple spatial and non-spatial processes, it is reasonable to assume that only some of these processes play a central role in representing “time in space” in the context of multitasking.
Study 2
In Study 2, we tested the hypothesis that individual and sex-related differences in multitasking reflect selective effects of spatial processing. Specifically, we relied on the notion that processing of spatial relations reflects two distinct aspects, often referred to as coordinate (metric) and categorical (relative) spatial processing (e.g., Huttenlocher, Hedges & Duncan, 1991; Kosslyn, 1987; Kosslyn et al., 1989; Newcombe & Huttenlocher, 2000). Categorical spatial relationships refer to equivalent classes of spatial positions relative to a perceptually distinguishable reference object (e.g., left/right, below/above, inside/outside). By contrast, coordinate spatial relationships refer to more precise spatial locations, which can be expressed in quantitative terms of (e.g., metric distances among spatial elements). Patient studies (e.g., Laeng, 1994, 2006; Palermo, Bureca, Matano, & Guariglia, 2008), experimental findings (Kosslyn, 1987; Kosslyn et al., 1989) and brain imaging studies (e.g., Baciu et al., 1999; Kosslyn, Thompson, Gitelman, & Alpert, 1998) support the distinction between categorical and coordinate spatial processing (for overviews see also Jager & Postma, 2003; Laeng et al., 2003; Voyer, Postma, Brake, & Imperato-McGinley, 2007).
Despite a general agreement on the coordinate versus categorical spatial processing distinction, sex differences in these specific processes have not been systematically investigated, and thus the empirical support for selective effects is non-existent. However, several lines of evidence would be consistent with such a difference. First, the distinction between categorical and coordinate processing is believed to reflect lateralization in the organizational structure of the brain, with several studies indicating that the left and right cerebral hemispheres may play different roles in processing these two spatial relations. Typically, processes supported by the left cerebral hemisphere show more proficiency in processing categorical spatial relations, whereas processes associated with the right cerebral hemisphere seem more effective in tasks based on coordinate spatial relations (see Hellige, Laeng, & Michimata, 2010; Jager & Postma, 2003; Laeng et al., 2003, for reviews). Considering that many cognitive sex-differences are believed to reflect similar differences in lateralization (see Halpern, 2012; Miller & Halpern, 2014; Vogel, Bowers & Vogel, 2003, for overviews), with males relying more on right-lateralized spatial processing compared to females, one might expect sex differences to be emphasized in coordinate spatial processing (but for evidence of sex differences in prefrontal lateralization see also Hugdahl, Thomsen & Ersland, 2006; Deutsch, Bourbon, Papanicolau, & Eisenberg, 1988).
These selective effects might also reflect sex differences in the use of spatial strategies, which in turn may reflect disparities in coordinate versus categorical processing. Studies on both animals (Suzuki, Augerinos & Black, 1980; Williams, Barnett & Meck, 1990) and humans (Dabbs, Chang, Strong & Milun, 1998; Saucier et al., 2002) point to differences in spatial orientation. Typically, males show preference for Euclidean properties of a room (i.e., more absolute coordinate processing), whereas females exhibit a preference for landmarks (i.e., more relative category-type of processing). Brain-imaging studies also provide some support for this line of reasoning. Hugdahl et al. (2006) reported a functional magnetic resonance imaging (fMRI) study showing sex differences in cortical (parietal vs. frontal) activation, during mental rotation and concluded that males are biased towards coordinate processing and females towards categorical processing (see also Jordan, Wüstenberg, Heinze, Peters & Jäncke, 2002; Rybash & Hoyer, 1992; Zacks, 2008).
In Study 2, all participants completed the counter task together with separate tasks of coordinate and categorical spatial processing. To this end, we used a modified version of the clock task (cf., Michimata, 1997; Palermo, Piccardi, Nori, Giusberti & Guariglia, 2012). As a reference measure of spatial processing, all participants also completed the MRT. To accentuate individual differences in spatial ability, female participants were either in the luteal phase of the menstrual cycle (during which their spatial ability is reduced) or in the menstrual phase of the cycle (during which sex differences in mental rotation are typically eliminated; Hausmann, Slabbekoorn, Van Goozen, Cohen-Kettenis & Güntürkün, 2000; Mäntylä, 2013). The primary prediction of Study 2 was that individual differences in multitasking reflect selective effects of coordinate (vs. categorical) spatial processing, and that these selective effects contribute to sex-related differences in multitasking.
A secondary aim of Study 2 was to relate individual differences in counter-task performance to a more realistic and validated task of multitasking. To this end, participants also completed the Simultaneous Capacity/Multitasking (SIMKAP) work simulation (Bratfisch & Hagman, 2003). The SIMKAP is composed of three main tasks that are performed simultaneously, along with a fourth task that periodically interrupts the other three tasks. These main tasks consist of simple operations such as crossing over matching numbers, doing word puzzles or remembering future intentions, and the fourth task resembled intentions that often occur via simulation of technology (e.g., responding to emails, looking up calendar schedules or retrieving phone numbers from an address book).
SIMKAP is a widely used commercial simulation of complex everyday multitasking with good measurement properties (Braun, Hüttges, Timm, Wieland & Willamowski, 2002; Bratfish & Hagman, 2003; Bühner, König, Pick & Krumm, 2006; der Horst, Klehe & Leeuwen, 2012). Compared to the counter task, individual differences in SIMKAP performance presumably reflect a wider variety of skills, including different kinds of spatial and nonspatial abilities, but the generality of our spatiotemporal hypothesis would be corroborated if (a) the two measures of multitasking correlated significantly, (b) showed similar sex differences, and (c) also SIMKAP performance was associated with spatial ability (in general terms due to its inherent complexity).
In Study 2, we also collected additional measures (albeit less relevant to the spatiotemporal hypothesis of multitasking), with the secondary aims of controlling for the role of additional potential factors (i.e., numeracy, the shifting facet of executive control). However, in order to prevent the negative consequences of fatigue, we had to limit the duration of the experimental session to a maximum of 2 hours. Thus it was not feasible to test all the participants on all these additional tasks. Therefore, we administered either the numeracy plus shifting measures or the SIMKAP task to two randomly selected subsets of participants. This strategy ensured that we achieved the main goal of Study 2 on the whole set of participants, yet allowing us to pursue two secondary aims (additional control and external validation) on two distinct subsets, while keeping the length of the testing session constant and hopefully limiting fatigue effects.
Method
Participants
A total of 122 university undergraduates (44 males) between 18 and 44 years of age (M = 25.6, SD = 5.6) participated in the study in return for partial course credit or a movie voucher. Sample size was determined with an a priori power analysis based on the correlation between MRT and counter task performance in Study 1 (estimated power .90).Footnote 2 As for the subgroup sizes, we followed our previous studies and recruited approximately 40 participants in each group (luteal, menstrual, and male).
The female participants completed the test session while being in either the menstrual phase (defined as 2–3 days before the predicted menstruation or during the first week of the cycle, 39 females), or in the luteal phase (defined as 2–3 days before ovulation or during the days of predicted ovulation, 37 females) of their menstrual cycle (see also Mäntylä, 2013). Individuals who were under hormonal treatment, used hormonal contraceptives, or were pregnant were not included in the study. Excessive computer gamers (>1.5 hours/day) were also excluded from the study. All participants were tested on the three spatial tasks, but participants who were tested on the SIMKAP (n = 42; 14 females from the menstrual group, 15 females from the luteal group and 13 men) did not complete the additional numeracy and switching tasks, and vice versa (n = 76; 25 females from the menstrual group, 22 females from the luteal group and 29 men).
Tasks and procedure
Spatial ability was assessed with three tasks, including the MRT. As in Study 1, participants completed two subsets (A and D) of the MRT, and the dependent measure of this task was the number of correct responses (max = 24 for both sets). The coordinate and categorical tasks were nominally identical in that participants were presented with digital time-readings (e.g., 07:10) on a computer screen, and were instructed to imagine the stimulus time as the hands of an imaginary analog clock. For the coordinate task, participants were asked to indicate which of two concurrently presented digital readings (e.g., 13:49 – 07:10) formed a larger angle between the hour and the minute hands on an imagined analog clock face (see Fig. 1, left panel). To equate task difficulty, the categorical task involved three concurrently presented digital readings (e.g., 13:49 – 07:10 – 02:37; see Fig. 1, right panel). Participants imagined the position of the clock hands relative to the four quadrants of an analog clock, and they indicated whether any of the quadrants were “free” (e.g., 13:49 and 07:10 occupy the first, third and fourth quadrants and 02:37 the first and fourth quadrants, respectively, leaving the second quadrant free). None of the stimuli resulted in vertical (6 and 12) or horizontal (3 and 9) hand positions that could be perceived as ambiguous. Both tasks comprised 20 items and the test phase was preceded by a set of practice items, during which an analog clock face (without the clock hands) was displayed as a support. In the coordinate task, the angular difference between a pair of time-readings was large (>60°) for half of the items and small (<60°) for the remaining pairs. Participants responded by pressing designated keys, and they were instructed to respond as quickly as possible while avoiding mistakes. Response time (max = 20 s) and accuracy were the dependent measures of both tasks. Spearman–Brown split-half reliability measure indicated very high internal consistency for the coordinate (α = .94) and categorical (α = .82) tasks.

A schematic illustration of the (a) coordinate and (b) categorical spatial tasks. The analog clock faces were presented only during the practice phase
Executive functioning was assessed with a computerized switching task (Monsell, 2003; Rogers & Monsell, 1995), in which the participants classified digits in terms of parity (odd vs. even) and magnitude (less or greater than 25). The separate conditions comprised 24 items and the alternate condition involved a set of 48 randomly mixed items. Participants responded by pressing designated keys, and they were instructed to respond as quickly as possible while avoiding mistakes. Switch cost in terms of difference in response time between the separate and alternate conditions was the dependent measure of the switching task. Individual differences in numeracy were assessed with the Swedish version of the Berlin Numeracy Test (BNT; Cokely, Galesic, Schulz, Ghazal & Garcia-Retamero, 2012).
Multitasking was measured with the counter task described in Study 1. We also used a more established and complex test, the SIMKAP, in order to assess the external validity of the counter task and further explore the relation between spatial ability and multitasking. SIMKAP is composed of three main parts, presented sequentially for 6 min each, with the first, number-crossing task, containing a multitude of numbers, where identical ones are to be crossed over using the computer mouse (see Fig. 2). In the second, letter-crossing, part the same rule is applied to letter combinations and in the third one, to geometrical figures. While continuously working on these tasks, participants are required to solve simple queries presented in the middle of the screen. In a field, at the bottom of the screen, 20 answers are presented, but only some of them are probable answers to the question. Again, by using the computer mouse, participants are asked to mark the correct answer. Also, presented through a headset, participants need to answer additional questions (e.g., “You are invited to lunch. On which day do you have time?”), the answers to which are to be looked up in a virtual calendar or a phone book, and some of them are to be answered with a delay (e.g., “When the timer shows 4:35, answer the following question …”). For the purpose, there is a timer running in the upper right corner of the screen and participants are allowed to write down, on a piece of paper, information about the answer and when it should be typed in. The component task measures (perceptual speed, number of correctly answered questions, number of correctly marked items, number of errors) are used for computing an overall measure of multitasking, which we used as the primary measure of SIMKAP performance.

A screenshot of SIMKAP showing all the different tasks presented simultaneously. Questions relating to the calendar of the phone book functions are presented through a headset
Each participant was tested individually during a 2-h session. Except for the MRT and the BNT, all the tasks were computerized and the stimuli were presented on a 20-in display. Each task included separate instructions and a practice phase during which the experimenter checked that instructions were properly understood. After completing a brief questionnaire about demographic background and computer-gaming habits, participants completed the tasks in a fixed order. Specifically, the order of the coordinate and counter tasks was counterbalanced and separated by the counter task session, and followed by the remaining cognitive tasks. Finally, before debriefing, all the female participants filled in a short questionnaire about their menstrual cycle and use of hormonal substances.
Results
As in Study 1, multitasking performance was based on a combined score of the four counter tasks, with response accuracy as the primary measure. The menstrual phase of three female participants could not be determined due to unclear self-reports, and one male participant did not complete the whole test session. Data of these participants were not included in the analysis (N = 118 for the complete data set). Five participants showed exceptionally fast response times (<3SD) in the coordinate or categorical tasks, and these data were excluded from the analyses.
Table 3 shows the correlations between the primary measures (the coordinate, categorical, and switching tasks were based on response time data). The three spatial tasks showed consistent and significant associations, and even more importantly, counter accuracy correlated significantly with all three measures. These findings provide additional support for the hypothesis that multitasking performance is related to individual differences in spatial ability. The correlation data also showed that both numeracy and task switching were related to counter performance (although for the latter correlation, p = .06).
Sex differences in spatial ability and multitasking
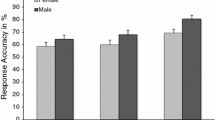
Figure 3 summarizes the main findings as a function of sex. These results suggest sex differences favoring men in most measures of multitasking and spatial ability. Analysis of the counter accuracy data showed a significant sex difference, F(1, 119) = 24.05, MSe = .02, η 2 = .18, p < .01, favoring males by about 10 % across the four counters. Furthermore, sex differences in multitasking were accentuated between males and females in the luteal phase of the menstrual cycle. An ANCOVA with numeracy and executive functioning (task switching) as covariates, confirmed this observation F(1, 60) = 9.08, MSe = .01, η 2 = .23, p < .01. Furthermore, post hoc comparisons showed a non-significant difference between males and females in the menstrual phase (p > .40). Separate analysis of the executive functioning and numeracy data showed no group differences (Fs < 1).

Sex differences in mental rotation (Mental Rotations Test, MRT), numeracy (Berlin Numeracy Test, BNT), coordinate and categorical task accuracy, and counter task accuracy. Error bars indicate standard error
A MANOVA on the spatial-task data (with MRT, coordinate, and categorical tasks as dependent measures) showed a significant main effect of sex, favoring men (Wilk’s Lambda = 0.76, F(3, 107) = 11.15, p < .01). Consistent with our hypothesis, separate ANOVAs on these data showed selective sex differences in spatial ability, with significant differences in MRT performance, F(1, 109) = 30.09, p < .01, and coordinate task accuracy F(1, 109) = 4.52, p < .05, but not in categorical task performance (F < 1).
As with the counter task data, these effects fluctuated across the female menstrual cycle, in that sex differences were accentuated for females in the luteal phase and eliminated for females tested at menses. Figure 4 illustrates this pattern of results, measured in terms of response time (the accuracy data showed the same pattern of results). An ANOVA on the coordinate task data showed a significant group effect, F(2, 115) = 4.51, MSe = 322962, η 2 = .09, p < .01, and post hoc tests indicated that the luteal group was significantly slower than males (p < .05). The mean of the menstrual group was not significantly different from those of the luteal group (p < .10) and males. As shown in Fig. 3, the three groups showed similar levels of categorical task performance (F < 1).

Mean response time as a function of task and group. Error bars indicate standard error
Selective effects of spatial processing on multitasking
Consistent with our hypothesis, participants with more efficient spatial abilities were better multitaskers than those with less efficient spatial skills, and these effects were accentuated by hormonal changes across the female menstrual cycle. To test the hypothesis that individual differences in multitasking performance reflect selective effects of spatial processing across sex, we completed a multiple regression analysis in which the MRT, coordinate and categorical tasks were used as predictors of counter performance. As expected, the MRT emerged as the greatest predictor of multitasking performance (Beta = .39, p < .01). More importantly, even when MRT was included in this analysis, coordinate (Beta = −.20, p < .01), but not categorical (Beta = −.10, p > .29) was a significant predictor of multitasking performance, R = .37, F(2, 109) = 8.8, MSe = .02, p < .01.Footnote 3 These findings proved to be robust even in a second multiple regression on the counter data, with numeracy, task switching, coordinate, and categorical scores as predictors.Footnote 4 This analysis indicated that coordinate processing (Beta = −.30, p < .05) and numeracy (Beta = .26, p = .05) were the main predictors of multitasking performance, R = .46, F(4, 58) = 3.96, MSe = .01, p < .01.
Thus, although the coordinate and categorical tasks were highly similar and equally difficult, coordinate processing was the primary predictor of multitasking performance, and this selective effect was observed even when individual differences in numeracy and executive function were taken into consideration.
Generality of findings: SIMKAP
As shown in Table 3, the SIMKAP and counter data were significantly correlated, suggesting reasonable external validity of the counter task. Consistent with the spatiotemporal hypothesis, also SIMKAP performance reflected spatial ability with significant correlations with the MRT (r = .39, p = .01), coordinate (r = .43, p < .01), and categorical (r = .47, p < .01) accuracy measures.Footnote 5 For the response-time data, shown in Table 3, the categorical, but not coordinate (p < .10), scores correlated with SIMKAP. Finally, the SIMKAP data showed similar sex difference as the counter task data in that males (mean = 6.25) outperformed females (mean = 4.87), F(1,40) = 4, MSe = .32, η 2 = .09, p < .05. A separate ANOVA with group (luteal, menstrual or male) as a between-subjects variable showed a marginal effect, with males having an advantage over the luteal females (mean = 4.29), F(1,38) = 3.11, MSe = .33, η 2 = .14, p = .06. No other effects were observed.
General discussion
The present findings suggest that spatial processing is important to multiple-task performance. In both our studies, involving individuals from different countries, participants with efficient spatial abilities were better multitaskers than individuals with more limited spatial skills. Extending these findings, a central contribution of this study was that individual differences in multitasking were selectively related to spatial relation processing. Individuals who were good at coordinate, rather than categorical, spatial processing were also better multitaskers than individuals with less efficient coordinate processing skills. This result is consistent with the notion that better multitasking performance is associated with a more precise (and demanding) transformation of temporal deadlines to a metric spatial representation, as compared to spatial processes relying on more relative spatial categories.
Another central finding of this study was that individual differences in both multitasking and spatial ability (in terms of mental rotation and coordinate processing) reflected sex- and hormone-related effects. Consistently, males were better multitaskers than females, and these effects were accentuated by hormonal changes across the female menstrual cycle. Specifically, sex differences in spatial ability were significant for women in the luteal phase, and eliminated for women tested at menses. This pattern of results suggests that coordinate processing, involving quantitative, rather than qualitative, judgments of spatial relations, is highly sensitive to fluctuations in sex steroids. By contrast, females in the luteal and menstrual phases of the cycle showed similar levels of performance on the categorical task, suggesting that processing of categorical spatial information is less sensitive to sex-hormone related effects.
Our findings are consistent with earlier studies on spatial orientation, suggesting that males rely more on quantitative spatial representations compared to females, who often solve the same spatial task by using more relative encoding strategies (see Coluccia & Louse, 2004, for a review). For example, Saucier et al. (2002) reported a study in which participants navigated to four unknown destinations by using either Euclidean-based or landmark-based instructions for directions. They found that males navigated best when provided with Euclidean information, whereas women performed best when provided with landmark information. Furthermore, females made more errors than males when using Euclidean-based instructions, whereas landmark-based instructions eliminated sex differences in navigation. As concluded by Coluccia and Louse (2004), these sex differences in orientation, in part, reflect differences in task difficulty, with landmark based (categorical) tasks being easier and less sensitive to sex differences than quantitative (coordinate) tasks, possibly due to differences in demands on visuospatial working memory load (e.g., Garden, Cornoldi & Logie, 2002; Pazzaglia, 1999). This result also fits well with the earlier suggestions that males rely more on right-lateralized spatial processing compared to females and that coordinate spatial processing is more right-lateralized than categorical spatial processing.
An important issue to be raised is the specific contribution of spatial processes to multitasking beyond executive functioning and visuo-spatial working memory (VSWM). Indeed, the presence of multiple digital stimuli in both the coordinate and categorical tasks, and the need to keep in mind visuo-spatial information while carrying out these tasks may require VSWM resources and related executive functions. However, our findings show that spatial abilities, in particular coordinate processing, matter for multitasking in addition to executive control and MRT, and this conclusion is strengthened by the fact that we employed executive tasks covering different facets of control in the present study. Also, converging findings on the specific role of spatial abilities have been obtained by using nonverbal executive control and working memory tasks (see Mäntylä, 2013; Todorov et al., 2014). Thus, the fact that coordinate processing contributed to multitasking performance beyond MRT in Study 2 is particularly interesting, because previous research showed that mental rotation processes place demands upon executive processes (Bruyer & Scailquin, 1998; Cornoldi & Mammarella, 2014; Salway & Logie, 1995) as well as involving VSWM, when keeping in mind the products of multiple rotation steps (Bauer & Jolicouer, 1996; Christophel, Cichy, Hebart & Haynes, 2015).
Consequently, even though all three spatial tasks used in this study involve other-than-spatial components, we extend earlier work by showing selective sex differences in spatial abilities beyond executive processes, related to specific demands on metric spatial representations. Moreover, these differences were detected despite the fact that both the coordinate and categorical tasks involved VSWM and related executive functions and, as noted earlier, the two tasks were equally difficult in terms of overall task accuracy. Furthermore, although some studies have shown sex differences in VSWM (Duff & Hampson, 2001; Geiger & Litwiller, 2005; Vecchi & Girelli, 1998), these differences do not explain the male superiority in mental rotation. For example, Kauffman (2007) found that VSWM mediated the relationship between sex and spatial ability, defined as a latent variable of visualization and mental rotation. However, there was also a direct effect of sex on the unique variance in mental rotation ability, and this effect was not mediated by working memory.
A pure working-memory-based explanation is also inconsistent with our hormone-related findings. Specifically, in contrast to mental rotation, many other types of cognitive functions exhibit either no relation to the cycle or even an improvement at high levels of estradiol (e.g., Hampson & Morley, 2013; Maki, Rich & Shayna Rosenbaum, 2002). For example, Hampson and Morley (2013) found that females with higher levels of circulating estradiol made significantly fewer errors on a spatial working memory task than males and female tested under low estradiol. Instead, the findings of Hampson, Levy-Coopermana and Korman (2014) suggest that the estradiol effect is related to transformation-specific computations required to carry out mental rotation, possibly located in the intraparietal sulcus (Zacks, 2008).
A methodological limitation of our study is that the assessment of menstrual phase in Study 2 was based on self-reports. Although these subjective measures strongly correlate with objective measures (e.g., Baker, Denning, Kostin & Scharwts, 1998), self-reports provide only a proxy for more specific sex-hormone-related effects on higher cognitive functions (see also Bean, Leeper, Wallace, Sherman & Jagger, 1979). Future studies should replicate our findings with hormone measurements, but it should be noted that these methodological limitations might actually have reduced (and not enhanced) the magnitude of our findings. Specifically, as the estimates of the luteal phase were indirect and probably less reliable than those of the menses, estradiol levels were not necessarily heightened for some of the female participants who reported being in the luteal phase at the time of testing. For example, Cantú, Simpson, Griskevicius, Weisberg, Durante and Beal (2013) reported that 30 % of the initially recruited participants were excluded from their study because they did not show heightened levels of sex hormones during the expected luteal phase.
In addition to sex- and hormone-related differences in both multitasking and spatial ability, the present findings suggest a more general association in that individuals with good performance in the coordinate task were also better multitaskers than individuals with less efficient coordinate task performance. Specifically, multiple regression analysis in Study 2 showed that coordinate (but not categorical) task performance was a significant specific spatial predictor of counter task performance, even when individual differences in task switching and numeracy were considered.
As noted earlier, a central assumption of the spatiotemporal hypothesis is that individual differences in spatial ability contribute to multitasking performance when the demands on temporal coordination are relatively high. These demands, in turn, are related to individual characteristics and task demands. In most everyday situations of multitasking (cf. preparing a breakfast; Rendell & Craik, 2000), overlearned scripts, schemas, many environmental cues and related knowledge structures reduce demands on task coordination by providing a spatiotemporal structure (a mental timeline) for goal-directed actions. Furthermore, even in conditions in which the support of these knowledge structures is reduced (as in our counter task), demands on temporal coordination can be low, for example, due to few component tasks, as in dual-task settings, or because the temporal constraints are rather flexible. Representing “time in space” under these conditions would not pose major task demands on task coordination, and therefore individual and sex-related differences in spatial ability should not mediate multitasking performance. As a support for this argument, sex differences are not typically observed in dual-task conditions (Strayer, Medeiros-Ward & Watson, 2013).
It might be reasonable to consider the possibility that stereotype threat (e.g., Steele, 1997; Steele & Aronson; 1995; see also Maass & Cadinu, 2003) may have contributed to gender differences in our study. Both the spatial tasks and the counter task (which is based on numbers) were relevant to the gender stereotype, and both the male and female participants might have been affected by these beliefs. However, our main findings highlighted very specific effects in terms of spatial ability, and thus it is quite unlikely that they were produced by a general influence of the stereotype threat. Moreover, some findings specifically militate against such assumption. First, participants were expecting a test of multitasking performance (along with “other” cognitive tasks). Considering the popular belief that women are better multitaskers than men, these gender stereotypes might actually have facilitated female performance. Second, as noted earlier, we observed selective sex differences (in both multitasking and spatial ability) between males and females in the luteal phase only. Furthermore, the difference between females in the luteal and menstrual phase was also selective in that it was observed in the coordinate, but not in the categorical, task (which were very similar and equally difficult).
Gender stereotype threats might have interacted with sex hormones in that only women in the luteal phase might have been sensitive to these effects. However, empirical support for this hypothesis is limited. For example, Halpern and Tan (2001; see also Hausmann, Schoofs, Rosenthal & Jordan, 2009) reported a study in which participants completed a mental-rotation task, a word-finding task, and a questionnaire concerning beliefs about females’ and males’ cognitive abilities. As expected, males outperformed females tested at the luteal phase, with the opposite sex difference in word finding. However, these sex-typical results could not be attributed to either stereotype threat (or more implicit stereotype activation) in that stereotypes about the cognitive abilities of males and females did not correspond to mental-rotation or word-finding performance.
The primary focus of our study was to examine individual differences in multiple-task performance when the demands on temporal coordination are relatively high. To this end, we used a rather simple and well-controlled, four-component counter task that captures essential aspects of multitasking (monitoring of multiple deadlines and switching between tasks), while attempting to eliminate the role of expertise and the unwanted influence of domain-specific and modality-specific subtasks requirements. However, in Study 2, we also examined the issue of external validity of our findings, by having some of the participants complete a more complex simulation of everyday multitasking (SIMKAP). Performance on the counter and SIMKAP tasks correlated significantly, and both tasks showed a significant correlation with the MRT. These correlations were less consistent for the coordinate and categorical spatial data, in that only the latter task showed a significant correlation with the SIMKAP scores, possibly due to the complexity of the SIMKAP and the multiple and varied requirements of its component subtasks. It should also noted that demands on planning and reliance on exact timelines are less pronounced in the SIMKAP than in the counter task. These findings suggest that the counter task engages partly similar processes as the SIMKAP and that spatial ability contributes to individual differences even in rather complex simulation of everyday multitasking.
In conclusion, the present findings suggest that spatial processing is important to multitasking scenarios with high demands on temporal coordination. In more general terms, the present findings may suggest that spatial abilities reflect a complementary function to executive control function by providing a form of mental scaffolding for complex temporal relations. Under conditions in which prior knowledge scripts and habitual schemas are not available or the demands on executive control are high due to task constraints and deadlines, spatial relation processing may facilitate multiple task monitoring and goal-directed behavior. By relying on patterns of mental timelines, demands on working memory and related executive control functions can be reduced and, as our findings suggest, a more fine-tuned metric representation of these spatiotemporal relations may provide a more accurate support than categorical relations. An interesting avenue for future work would be to examine the relative importance of executive control functions, spatial abilities, and prior knowledge on goal-directed behavior in different contexts of everyday multitasking.
Notes
To control for potential differences in the arrangement of the colored occluders on the computer screen, we used two versions of the counter task. As expected, the two versions showed the same pattern of results, and thus the data were pooled.
Specifically, in Study 1 we found a correlation of .297 between MRT and the counter task (valid n = 99), and a post-hoc power analysis showed a good power (.85, two tailed). Based on an estimated minimum correlation of .30 between MRT and counter, we then estimated the approximate number of participants for the power of .90. An a-priori power analysis for Study 2 provided an estimate of n = 112 for a .90 power (two tailed). As a consequence, we selected 122 participants as the initial sample size for Study 2, assuming a potential loss of participants during the study.
We conceive MRT as a higher-level measure of spatial ability (see also Kaufman, 2005) that possibly involves both categorical and coordinate processing, albeit to a different extent (Carpenter, 1985). Therefore, including it in the multiple regressions might introduce a source of theoretical confusion and it might complicate the interpretation of the findings on the relative role of categorical vs. coordinate processing in multitasking. Moreover, the two more basic spatial tasks were equated for difficulty independently of the MRT, and this might represent an additional confound.
This analysis was carried out on the task switching and numeracy measures, thus on a reduced set of participants. Starting from the correlations between the counter task and task switching (-.21) and BNT (.27*), we estimated power with a post-hoc power analysis, obtaining two-tailed estimates of .43 and .64, respectively. Overall, these analyses show that the power of our tests was rather low, and thus suggest some caution in the interpretation of the results. However, the findings are consistent with Study 1 results, our previous studies (e.g., Mäntylä, 2013), and with the main findings of Study 2, and this provides strong converging evidence for our conclusions on the specific role of spatial abilities in the counter task.
For the SIMKAP, a post-hoc power analysis carried out on the SIMKAP-counter correlation (r = .48) showed a very good power (.93), and the SIMKAP-MRT correlation (.39) was associated with a reasonable good power (.78). Therefore, these analyses do not expose particular power problems with the SIMKAP-related tests.
References
Baciu, M., Koenig, O., Vernier, M. P., Bedoin, N., Rubin, C., & Segebarth, C. (1999). Categorical and coordinate spatial relations: fMRI evidence for hemispheric specialization. NeuroReport, 10, 1373–1378.
Baker, A. H., Denning, A. C., Kostin, I., & Schwartz, L. (1998). How accurate are women's estimates of date of onset of next menses? Psychology and Health, 13, 897–908.
Bauer, B., & Jolicoeur, P. (1996). Stimulus dimensionality effects in mental rotation. Journal of Experimental Psychology: Human Perception and Performance, 22, 82.
Bean, J. A., Leeper, J. D., Wallace, R. B., Sherman, B. M., & Jagger, H. (1979). Variations in the reporting of menstrual histories. American Journal of Epidemiology, 109, 181–185.
Bonato, M., Zorzi, M., & Umiltà, C. (2012). When time is space: Evidence for a mental time line. Neuroscience and Biobehavioral Reviews, 36, 2257–2273.
Bratfisch, O., & Hagman, E. (2003). Simultankapazität/Multi-Tasking (SIMKAP) Version 24.00: Handanweisung (Simultaneous capacity/multi-tasking (SIMKAP) release 24.00: Manual). Mödling, Austria: Schuhfried.
Braun, V., Hüttges, M., Timm, E., Wieland, R., & Willamowski, A. (2002). Instrumente zur Personalauswahl und Personalentwicklung im Call Center (CCall report 13)[Instruments for personnel selection and development in call centres (CCall report 13)]. Hamburg, Germany: Verwaltungs-Berufsgenossenschaft.
Bruyer, R., & Scailquin, J. C. (1998). The visuospatial sketchpad for mental images: Testing the multicomponent model of working memory. Acta Psychologica, 98, 17–36.
Bühner, M., König, C. J., Pick, M., & Krumm, S. (2006). Working memory dimensions as differential predictors of the speed and error aspect of multitasking performance. Human Performance, 19, 253–275.
Burgess, P. W., Veitch, E., de Lacy Costello, A., & Shallice, T. (2000). The cognitive and neuroanatomical correlates of multitasking. Neuropsychologia, 38, 848–863.
Cantú, S. M., Simpson, J. A., Griskevicius, V., Weisberg, Y. J., Durante, K. M., & Beal, D. J. (2013). Fertile and selectively flirty women’s behavior toward men changes across the ovulatory cycle. Psychological Science, 25, 431–438.
Casasanto, D., & Boroditsky, L. (2008). Time in the mind: Using space to think about time. Cognition, 106, 579–593.
Ceci, S. J., & Bronfenbrenner, U. (1985). “Don't forget to take the cupcakes out of the oven”: Prospective memory, strategic time-monitoring, and context. Child Development, 56, 152–164.
Christophel, T. B., Cichy, R. M., Hebart, M. N., & Haynes, J. D. (2015). Parietal and early visual cortices encode working memory content across mental transformations. NeuroImage, 106, 198–206.
Cokely, E. T., Galesic, M., Schulz, E., Ghazal, S., & Garcia-Retamero, R. (2012). Measuring risk literacy: The Berlin numeracy test. Judgment and Decision Making, 7, 25–47.
Coluccia, E., & Louse, G. (2004). Gender differences in spatial orientation: A review. Journal of Environmental Psychology, 24, 329–340.
Cornoldi, C., & Mammarella, I. C. (2014). 6 The organization of visuospatial working memory. Spatial Working Memory, 102.
Dabbs, J. M., Jr., Chang, E. L., Strong, R. A., & Milun, R. (1998). Spatial ability, navigation strategy, and geographic knowledge among men and women. Evolution and Human Behavior, 19, 89–98.
Dehaene, S., & Brannon, E. M. (2011). Space, time and number in the brain: Searching for the foundations of Mathematical Thought. London: Elsevier.
Del Missier, F., Mäntylä, T., & Bruine de Bruin, W. (2012). Decision-making competence, executive functioning, and general cognitive abilities. Journal of Behavioral Decision Making, 25, 331–351.
der Horst, A. C., Klehe, U. C., & Leeuwen, L. (2012). Doing it all at once: Multitasking as a predictor of call center agents’ performance and performance-based dismissal. International Journal of Selection and Assessment, 20, 434–441.
Deutsch, G., Bourbon, W. T., Papanicolaou, A. C., & Eisenberg, H. M. (1988). Visuospatial tasks compared via activation of regional cerebral blood flow. Neuropsychologia, 26, 445–452.
Duff, S. J., & Hampson, E. (2001). A sex difference on a novel spatial working memory task in humans. Brain and Cognition, 47, 470–493.
Fisher, H. (1999). The first sex: The natural talents of women and how they are changing the world. New York, NY: Random House.
Fuster, J. M. (1993). Frontal lobes. Current Opinion in Neurobiology, 3, 160–165.
Garden, S., Cornoldi, C., & Logie, R. H. (2002). Visuo-spatial working memory in navigation. Applied Cognitive Psychology, 16, 35–50.
Geiger, J. F., & Litwiller, R. M. (2005). Spatial working memory and gender differences in science. Journal of Instructional Psychology, 32, 49–57.
Halpern, D. F. (2012). Sex differences in cognitive abilities (4th ed.). New York, NY: Psychology Press.
Halpern, D. F., & Tan, U. (2001). Stereotypes and steroids: Using a psychobiosocial model to understand cognitive sex differences. Brain and Cognition, 45, 392–414.
Hampson, E., Levy-Cooperman, N. A., & Korman, J. M. (2014). Estradiol and mental rotation: Relation to dimensionality, difficulty, or angular disparity? Hormones and Behavior, 65, 238–248.
Hampson, E., & Morley, E. E. (2013). Estradiol concentrations and working memory performance in women of reproductive age. Psychoneuroendocrinology, 38, 2897–2904.
Hausmann, M., Schoofs, D., Rosenthal, H. E., & Jordan, K. (2009). Interactive effects of sex hormones and gender stereotypes on cognitive sex differences—A psychobiosocial approach. Psychoneuroendocrinology, 34, 389–401.
Hausmann, M., Slabbekoorn, D., Van Goozen, S. H., Cohen-Kettenis, P. T., & Güntürkün, O. (2000). Sex hormones affect spatial abilities during the menstrual cycle. Behavioral Neuroscience, 114, 1245.
Hellige, J. B., Laeng, B., & Michimata, C. (2010). Processing asymmetries in the visual system. In K. Hugdahl (Ed.), The two halves of the brain (pp. 379–415). Cambridge, MA: MIT Press.
Hugdahl, K., Thomsen, T., & Ersland, L. (2006). Sex differences in visuo-spatial processing: An fMRI study of mental rotation. Neuropsychologia, 44, 1575–1583.
Huttenlocher, J., Hedges, L. V., & Duncan, S. (1991). Categories and particulars: Prototype effects in estimating spatial location. Psychological Review, 98, 352–376.
Jager, G., & Postma, A. (2003). On the hemispheric specialization for categorical and coordinate spatial relations: A review of the current evidence. Neuropsychologia, 41, 504–515.
Jordan, K., Wüstenberg, T., Heinze, H. J., Peters, M., & Jäncke, L. (2002). Women and men exhibit different cortical activation patterns during mental rotation tasks. Neuropsychologia, 40, 2397–2408.
Kauffman, S. B. (2007). Sex differences in mental rotation and spatial visualization ability: Can they be accounted for by differences in working memory capacity? Intelligence, 35, 211–223.
Kosslyn, S. M. (1987). Seeing and imagining in the cerebral hemispheres: A computational approach. Psychological Review, 94, 148–175.
Kosslyn, S. M., Koenig, O., Barrett, A., Cave, C. B., Tang, J., & Gabrieli, J. D. (1989). Evidence for two types of spatial representations: Hemispheric specialization for categorical and coordinate relations. Journal of Experimental Psychology: Human Perception and Performance, 15, 723–735.
Kosslyn, S. M., Thompson, W. L., Gitelman, D. R., & Alpert, N. M. (1998). Neural systems that encode categorical versus coordinate spatial relations: PET investigations. Psychobiology, 26, 333–347.
Laeng, B. (1994). Lateralization of categorical and coordinate spatial functions: A study of unilateral stroke patients. Journal of Cognitive Neuroscience, 6, 189–203.
Laeng, B. (2006). Constructional apraxia after left or right unilateral stroke. Neuropsychologia, 44, 1595–1606.
Laeng, B., Chabris, C. F., & Kosslyn, S. M. (2003). Asymmetries in encoding spatial relations. In R. Davidson & K. Hugdahl (Eds.), The asymmetrical brain (pp. 303–339). Cambridge, MA: The MIT Press.
Logie, R. H., Trawley, S., & Law, A. S. (2011). Multitasking: Multiple, domain-specific cognitive functions in a virtual environment. Memory & Cognition, 39, 1561–1574.
Maass, A., & Cadinu, M. (2003). Stereotype threat: When minority members underperform. European Review of Social Psychology, 14, 243–275.
Maki, P. M., Rich, J. B., & Shayna Rosenbaum, R. (2002). Implicit memory varies across the menstrual cycle: Estrogen effects in young women. Neuropsychologia, 40, 518–529.
Mäntylä, T. (2013). Gender differences in multitasking reflect spatial ability. Psychological Science, 24, 514–520.
Mäntylä, T., Carelli, M. G., & Forman, H. (2007). Time monitoring and executive functioning in children and adults. Journal of Experimental Child Psychology, 96, 1–19.
Mäntylä, T., Del Missier, F., & Nilsson, L. G. (2009). Age differences in multiple measures of time-based prospective memory. Aging, Neuropsychology, and Cognition, 16, 708–720.
Mäntylä, T., & Todorov, I. (2013). Questioning anecdotal beliefs and scientific findings: A reply to Strayer, Medeiros-Ward, and Watson (2013). Psychological Science, 24, 811–812.
Meyer, D. E., & Kieras, D. E. (1997). A computational theory of executive cognitive processes and multiple-task performance: Part 1. Basic mechanisms. Psychological Review, 104, 3–65.
Michimata, C. (1997). Hemispheric processing of categorical and coordinate spatial relations in vision and visual imagery. Brain and Cognition, 33, 370–387.
Miller, D. I., & Halpern, D. F. (2014). The new science of cognitive sex differences. Trends in Cognitive Sciences, 18, 37–45.
Miyake, A., Friedman, N. P., Emerson, M. J., Witzki, A. H., Howerter, A., & Wager, T. D. (2000). The unity and diversity of executive functions and their contributions to complex “frontal lobe” tasks: A latent variable analysis. Cognitive Psychology, 41, 49–100.
Monsell, S. (2003). Task switching. Trends in Cognitive Sciences, 7, 134–140.
Navon, D., & Gopher, D. (1979). On the economy of the human-processing system. Psychological Review, 86, 214.
Newcombe, N. S., & Huttenlocher, J. (2000). Making space: The development of spatial representation and spatial reasoning. Cambridge, MA: MIT Press.
Norman, D. A., & Bobrow, D. G. (1975). On data-limited and resource-limited processes. Cognitive Psychology, 7, 44–64.
Palermo, L., Bureca, I., Matano, A., & Guariglia, C. (2008). Hemispheric contribution to categorical and coordinate representational processes: A study on brain-damaged patients. Neuropsychologia, 46, 2802–2807.
Palermo, L., Piccardi, L., Nori, R., Giusberti, F., & Guariglia, C. (2012). The roles of categorical and coordinate spatial relations in recognizing buildings. Attention, Perception, & Psychophysics, 74, 1732–1741.
Pashler, H., & Wagenmakers, E. J. (2012). Editors’ introduction to the special section on replicability in psychological science a crisis of confidence? Perspectives on Psychological Science, 7, 528–530.
Pazzaglia, F. (1999). The role of distinct components of visuo-spatial working memory in the processing of texts. Memory, 7, 19–41.
Pease, A., & Pease, B. (2003). Why men can only do one thing at a time and women never stop talking. Bhopal, India: Orion.
Peters, M., Laeng, B., Latham, K., Jackson, M., Zaiyouna, R., & Richardson, C. (1995). A redrawn Vandenberg and Kuse mental rotations test-different versions and factors that affect performance. Brain and Cognition, 28, 39–58.
Rendell, P. G., & Craik, F. I. (2000). Virtual week and actual week: Age‐related differences in prospective memory. Applied Cognitive Psychology, 14, S43–S62.
Rogers, R. D., & Monsell, S. (1995). Costs of a predictible switch between simple cognitive tasks. Journal of Experimental Psychology: General, 124, 207.
Rybash, J. M., & Hoyer, W. J. (1992). Hemispheric specialization for categorical and coordinate spatial representations: A reappraisal. Memory & Cognition, 20, 271–276.
Salvucci, D. D., & Taatgen, N. A. (2008). Threaded cognition: An integrated theory of concurrent multitasking. Psychological Review, 115, 101–130.
Salway, A. F., & Logie, R. H. (1995). Visuospatial working memory, movement control and executive demands. British Journal of Psychology, 86, 253–269.
Saucier, D. M., Green, S. M., Leason, J., MacFadden, A., Bell, S., & Elias, L. J. (2002). Are sex differences in navigation caused by sexually dimorphic strategies or by differences in the ability to use the strategies? Behavioral Neuroscience, 116, 403.
Shallice, T., & Burgess, P. (1991). Deficits in strategy application following frontal lobe damage in man. Brain, 114, 727–741.
Steele, C. M. (1997). A threat in the air: How stereotypes shape intellectual identity and performance. American Psychologist, 52, 613.
Steele, C. M., & Aronson, J. (1995). Stereotype threat and the intellectual test performance of African Americans. Journal of Personality and Social Psychology, 69, 797.
Strayer, D. L., Medeiros-Ward, N., & Watson, J. M. (2013). Gender Invariance in Multitasking A Comment on Mäntylä (2013). Psychological Science, 24, 809–810.
Suzuki, S., Augerinos, G., & Black, A. H. (1980). Stimulus control of spatial behavior on the eight-arm maze in rats. Learning and Motivation, 11, 1–18.
Todorov, I., Del Missier, F., & Mäntylä, T. (2014). Age-related differences in multiple task monitoring. PLoS One, 9, e107619.
Vandenberg, S. G., & Kuse, A. R. (1978). Mental rotations, a group test of three-dimensional spatial visualization. Perceptual and Motor Skills, 47, 599–604.
Vecchi, T., & Girelli, L. (1998). Gender differences in visuo-spatial processing: The importance of distinguishing between passive storage and active manipulation. Acta Psychologica, 99, 1–16.
Vogel, J. J., Bowers, C. A., & Vogel, D. S. (2003). Cerebral lateralization of spatial abilities: A meta-analysis. Brain and Cognition, 52, 197–204.
Voyer, D., Postma, A., Brake, B., & Imperato-McGinley, J. (2007). Gender differences in object location memory: A meta-analysis. Psychonomic Bulletin & Review, 14, 23–38.
Watson, J. M., & Strayer, D. L. (2010). Supertaskers: Profiles in extraordinary multitasking ability. Psychonomic Bulletin & Review, 17, 479–485.
Wickens, C. D. (2002). Multiple resources and performance prediction. Theoretical Issues in Ergonomics Science, 3, 159–177.
Williams, C. L., Barnett, A. M., & Meck, W. H. (1990). Organizational effects of early gonadal secretions on sexual differentiation in spatial memory. Behavioral Neuroscience, 104, 84–97.
Zacks, J. M. (2008). Neuroimaging studies of mental rotation: A meta-analysis and review. Journal of Cognitive Neuroscience, 20, 1–19.
Acknowledgments
We thank Chiara Parma for collecting and scoring the data for Study 1, and Lars Jonasson and Jelena Corovic for their help with the data collection for Study 2. We would also like to thank three anonymous reviewers of the paper for their valuable input on an earlier version of the manuscript. This research was supported by the Swedish Research Council.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
About this article

Cite this article
Todorov, I., Del Missier, F., Konke, L.A. et al. Deadlines in space: Selective effects of coordinate spatial processing in multitasking. Mem Cogn 43, 1216–1228 (2015). https://doi.org/10.3758/s13421-015-0529-z
Published:
Issue Date:
DOI: https://doi.org/10.3758/s13421-015-0529-z