
Abstract
Contextual similarity between learning and test phase has been shown to be beneficial for memory retrieval. Negative priming is known to be caused by multiple processes; one of which is episodic retrieval. Therefore, the contextual similarity of prime and probe presentations should influence the size of the negative priming effect. This has been shown for the visual modality. In Experiment 1, an auditory four-alternative forced choice reaction time task was used to test the influence of prime-probe contextual similarity on negative priming and the processes underlying the modulation by context. The negative priming effect was larger when the auditory context was repeated than when it was changed from prime to probe. The modulation by context was exclusively caused by an increase in prime response retrieval errors in ignored repetition trials with context repetition, whereas repeating only the context but not the prime distractor did not lead to an increase in prime response retrieval. This exact pattern of results was replicated in Experiment 2. The findings suggest that contextual information is integrated with prime distractor and response information. Retrieval of the previous episode, including prime distractor, prime response, and context (event file), can be triggered when the former prime distractor is repeated, whereas a context cue alone does not retrieve the event file. This suggests an event file structure that is more complicated than its usually assumed binary structure.
Similar content being viewed by others

There is a long and ongoing research interest in the determinants of the negative priming effect, a phenomenon that has been extensively studied to understand the consequences of ignoring information (for a recent review, see Frings, Schneider, & Fox, 2015). Originally, the slowed-down responding to a stimulus previously used as a distractor was interpreted as the residue of an inhibitory process applied to the abstract representation of this previously to-be-ignored stimulus (Tipper, 1985). However, several alternative explanations have been proposed since (Milliken, Joordens, Merikle, & Seiffert, 1998; Neill & Valdes, 1992; Neill, Valdes, Terry, & Gorfein, 1992; Park & Kanwisher, 1994). Most prominently, negative priming has been conceptualized as a memory phenomenon. Based on the instance theory of automatization (Logan, 1988), which assumes that each encounter with a stimulus—a so-called instance—is encoded in memory and that the repetition of a stimulus leads to automatic retrieval of previous instances of this stimulus, Neill and Valdes (1992) postulated the episodic retrieval account. This account assumes that responding in each trial of a sequential response task is either accomplished by algorithmically applying the task rule to generate the correct response or by retrieving the response from memory, with the faster of the two processes determining the response. It is further assumed that the to-be-ignored prime stimulus is tagged with a “do-not-respond” flag. When the tagged stimulus reappears as the probe target, the corresponding instance from the prime is automatically retrieved which results in a conflict between the retrieved “do-not-respond” tag and the need to respond to the stimulus in the current trial. Responding in these so-called ignored repetition probe trials is typically slowed-down compared with control trials devoid of any repetitions between successive prime and probe presentations. A variant of the episodic retrieval account has been proposed by Rothermund, Wentura, and De Houwer (2005) and Mayr and Buchner (2006). Instead of assuming that a “do-not-respond” information associated with the prime distractor is retrieved in the probe, these authors postulated that information about the actually executed prime response is associated with the prime distractor and retrieved when the prime distractor is repeated as the probe target (referred to as prime–response retrieval variant of the episodic retrieval account, cf. Mayr & Buchner, 2006, p. 937, or stimulus–response retrieval account, cf. Rothermund et al., 2005, p. 482). In ignored repetition trials, the required prime and probe responses differ so that the retrieval of the previous prime response is inappropriate for probe responding, thereby impairing probe response speed and/or accuracy. In line with the assumption of prime response retrieval it has been shown repeatedly that participants commit more probe errors using the former prime response in ignored repetition than in control trials (Mayr & Buchner, 2006, 2010a, 2010b, 2014; Mayr, Buchner, & Dentale, 2009a; Mayr, Buchner, Möller, & Hauke, 2011a; Mayr, Hauke, & Buchner, 2009b; Mayr, Möller, & Buchner, 2011b). This specific finding is neither predicted by the nonresponse variant of the episodic retrieval account nor by an inhibitory account of negative priming.
Note that the negative priming phenomenon can also be interpreted against the background of the event-file framework (Hommel, 1998, for an overview see Hommel, 2004). Following this framework, features of stimuli as well as responses are linked together into common representations, so-called stimulus-response episodes or event files. The whole content of an event file is retrieved whenever individual features are reencountered. Applied to the negative priming phenomenon, the observed increase in prime response errors when the prime distractor is repeated as the probe target (Mayr & Buchner, 2006) suggests that prime distractor and prime response were bound together during prime processing into an event file. This event file is assumed to be fully retrieved by reencountering one of its components.
The different accounts that have been proposed to explain negative priming could all be corroborated by empirical evidence, leading to the conclusion that the negative priming phenomenon is the result of multiple processes (Frings et al., 2015, see also D’Angelo, Thomson, Tipper, & Milliken, 2016). The separation and understanding of the processes that underlie negative priming, as well as the investigation of how context affects these processes are the central aims of the present study.
We will first focus on the role of context on negative priming and then point out how to separate and pinpoint the different processes that underly negative priming and its modulation by context.
The role of context in negative priming
We define context as all sensations—either external (e.g., auditory or visual) or internal (e.g., specific emotional or physiological state)—that appear or are present when the task-associated stimuli (target and distractor stimuli in the case of the negative priming paradigm) are presented but that remain response-irrelevant throughout the whole task.
The so-called encoding specificity hypothesis (Tulving, 1983) implies that contextual information encoded during learning serves as an effective retrieval cue when it is later repeated. Accordingly, the contextual similarity between encoding and retrieval situations has been shown to be beneficial for successful retrieval (for a review and meta-analysis of context-dependent memory research, see Smith & Vela, 2001).
Assuming that an episodic memory retrieval process contributes to the negative priming effect with the prime display regarded as the learning phase and the probe display regarded as the retrieval phase, increased contextual prime-probe similarity should increase the negative priming effect. More specifically, when the prime context is repeated in the probe display, then the context should function as a retrieval cue that increases the probability of retrieving the former prime episode in ignored repetition trials. The increased probability of retrieving prime episode information, in turn, should increase the probability of a conflict with the current response requirement due to the retrieval of either non-response information (Neill & Valdes, 1992) or prime-response information (Mayr & Buchner, 2006; Rothermund et al., 2005). In contrast, the notion that negative priming is the after-effect of an inhibitory process is difficult to reconcile with a modulatory effect of prime-probe contextual similarity on negative priming. Inhibition of the distractor stimulus—be it the inhibition of an abstract representation or the inhibition of the individual features of a highly specific representation (the latter following the notion of Houghton & Tipper, 1994)—is thought to be established in the prime and to endure to the subsequent probe presentation in which the residue of the suppression impedes the processing of this representation and, hence, responding. Therefore, a forward-acting inhibition of the distractor representation should depend on the conditions during initial selection in the prime presentation but not on the match between prime and probe.
Several studies investigated the effect of prime-probe contextual similarity on negative priming in the visual modality. However, some of these studies (Fox & de Fockert, 1998; Stolz & Neely, 2001; Tse, Hutchison, & Li, 2011) confounded prime-probe contextual similarity with exact stimulus repetition (for details, see Tse et al., 2011). Studies without the confound showed that the negative priming effect increased when the prime and probe context were the same relative to a situation in which both contexts differed (Chao, 2009; Chao & Yeh, 2008; Neill, 1997; but see Wong, 2000). The modulatory effect of context repetition has been shown to be sensitive to temporal presentation parameters (see Experiment 1 by Neill, 1997, compared with Experiment 2 by Moore, 1996), spatial configurations (Chao & Yeh, 2008), and the variability of contextual cues (Chao, 2009).
The role of context has also been brought up recently in studies on stimulus-response binding and retrieval (Frings, Koch, & Moeller, 2017; Frings & Rothermund, 2017). For example, Frings and Rothermund (2017) investigated for the visual modality whether contextual features (such as color) are integrated into event files. They found a facilitative effect of context color repetition on responding if the response repeated between prime and probe (as compared to response change trials), which was interpreted as evidence of the integration of context and response features into a common event file. Interestingly, perceptual principles, such as figure-ground segmentation, modulated this integration process as only figural elements of the context (such as color presented in a small and clearly confined area) were integrated but not background elements of the context (such as a homogeneously colored entire screen).
Although there is evidence from negative priming and stimulus-response binding tasks that context can function as an effective retrieval cue and is integrated into stimulus-response episodes under certain conditions, the existing evidence is confined to the visual modality. Whereas in vision head and eye movements might be used to focus on task-related information, the omnipresence of sound and the lack of any peripheral mechanisms to shift the auditory focus might result in different and possibly stronger context effects in the auditory domain. Note that research from classical paradigms using recall, recognition, or cued recall tests to probe memory is ambiguous with respect to the role of modality in context-dependent memory. In the meta-analysis by Smith and Vela (2001), studies were differentiated by stimulus presentation mode (visual vs. auditory with respect to the target information). While context reinstatement (i.e., context repetition between learning and test phase) effects were descriptively larger for the auditory than for the visual modality, the difference in effect sizes was not significant.
Pinpointing the processes underlying the negative priming effect
With the exception of Tse et al. (2011)Footnote 1 none of the studies investigating the effect of contextual similarity on negative priming was designed to separate the different processes underlying the effect. Our aim in the present study was to test whether prime response retrieval processes as opposed to episodic retrieval processes or inhibitory processes underlie the assumed modulation of negative priming by contextual similarity. According to the prime response retrieval variant of the episodic retrieval model, the retrieval of prime response information triggered by the repetition of the former prime distractor stimulus during the probe should increase probe response errors by selectively increasing the probability with which the previous prime response is erroneously executed in ignored repetition trials as compared with control trials.
The four-alternative forced choice reaction time task employed in the present study has been successfully used to measure the contribution of prime response retrieval processes to the negative priming effect (Mayr & Buchner, 2006, 2010a, 2010b, 2014; Mayr, Buchner, et al., 2009b; Mayr, Buchner, et al., 2011a; Mayr, Hauke, et al., 2009a; Mayr, Möller, et al., 2011b). With this task, prime response errors can be separated from other types of errors (such as erroneous responses to the probe distractor). By contrast, in standard two-alternative categorization tasks, it is not possible to separate prime response errors from erroneous responses to the probe distractor, because the prime target and the probe distractor usually belong to the same response category. Based on the extent to which prime response errors are committed, the strength of a prime response retrieval process can be estimated and compared between ignored repetition and control trials using a multinomial measurement model (Mayr & Buchner, 2006, for details, see further below).
We used a multistaged testing strategy in the present experiments. We first tested whether contextual similarity effects on auditory negative priming are parallel to or differ from those reported for the visual modality. The negative priming effect should be larger when the context repeats between prime and probe than when it changes. Such an increase in effect size could become evident in reaction times, response errors, or both. In the second step, the effect of contextual similarity was analyzed in more detail. To this end, we analyzed the involvement of prime response retrieval processes. Specifically, we analyzed contextual similarity effects on prime response errors in ignored repetition compared with control trials, expecting a larger increase in prime response errors in ignored repetition trials when the context is repeated than when it is changed. Finally, we analyzed processes other than prime response retrieval that may be involved in generating the negative priming effect. For this purpose, we calculated adjusted error rates by excluding all prime response retrieval errors and by analyzing whether the adjusted error rates were sensitive to the contextual similarity manipulation.Footnote 2
To anticipate the results of Experiment 1, we found that the negative priming effect increases with increasing contextual similarity and that prime response retrieval processes underlie this modulation by context. To assess the reliability of these findings, we replicated the study in Experiment 2 at a different location with a different setup. We will report Experiment 1 and Experiment 2 in direct succession before we discuss the findings.
Experiment 1
Method
Participants
Participants were 58 adults (24 males), most of whom were students at Heinrich Heine University Düsseldorf. Their age ranged from 20 to 37 years (M = 24). Participants received partial course credit or monetary compensation for their participation. All participants reported normal hearing. Data sets of nine further participants had to be excluded. Eight participants were tested while a humming malfunctioning air-conditioning was running in the testing room. One participant committed more than 50% erroneous responses in several experimental conditions (compared with an average of approximately 9% errors in all relevant experimental conditions), which strongly suggests either a misunderstanding of the task or a lack of willingness to follow the instructions. Three further participants did not pass the training criterion (see below).
Materials and Task
Materials, task, and procedure were similar to preceding experiments that used the same experimental task (Mayr & Buchner, 2014). Four environmental sounds (frog, piano, drum, and bell) were used, each of which was 300-ms long. The sounds were played over headphones that were plugged into an Apple iMac computer which controlled the experiment. The sounds had an average loudness of approximately 70 db(A), loudness measurements (LA eq) were made at one side of the headphone while the four sounds were played in a loop.
In each presentation, two sounds were played simultaneously: one to the left and one to the right ear. While one sound had to be attended (target), the other had to be ignored (distractor). A 20-ms metronome click that preceded the sound pair presentation indicated the ear at which the to-be-attended sound would be played. Participants responded to the attended sound by pressing a response button assigned to this sound. Response buttons were four buttons on an external USB-connected response box. The response buttons were aligned sagittally to minimize spatial compatibility effects between the sounds’ location (left or right ear) and the required response. A total of 31 participants had to press the two distal buttons (frog and piano) with the middle and index fingers of their right hands and the two proximal buttons (drum and bell) with the middle and index fingers of their left hands. For the remaining 27 participants, the arrangement was reversed.
Simultaneously with each sound pair presentation, a context tone was played, which could either be a 300-Hz sine tone or a 700-Hz sine tone. The context tones were of the same duration as the target and distractor sounds (300 ms), including attack and decay intervals of 10-ms each. The context tones were played binaurally, so that they seemed to come from the center (and neither from the left or right side). They were easily distinguishable from the target and distractor sounds but also from each other. When added to the target and distractor sound pair, the overall loudness increased only moderately (<1 db(A) for the 300-Hz context tone and by approximately 5 db(A) for the 700-Hz context tone).
Experimental trials comprised a prime and a probe presentation. In ignored repetition trials, the ignored prime was identical to the attended probe with no other stimulus repetitions. For each ignored repetition trial, a parallel control trial was constructed by replacing the ignored prime with the remaining fourth stimulus. If only ignored repetition and control trials had been implemented, participants would have learned that the prime target and response never repeated as probe target and response. Therefore, the experiment also comprised attended repetition trials with a prime-to-probe repetition of the target stimulus and response but with no other stimulus repetitions. For each attended repetition trial, a parallel attended repetition control trial was constructed by replacing the attended prime with the remaining fourth stimulus (see Table 1 for examples of the different trial types). The results of the attended repetition and attended repetition control trials are not reported in the main text, because there were no hypothesis concerning performance in these trial types (but see the Appendix for a description and discussion of the results in the Attended Repetition and Attended Repetition Control trials).
The basic set of experimental trials comprised 48 trials: 12 ignored repetition, 12 control, 12 attended repetition, and 12 attended repetition control trials.Footnote 3 This basic set was implemented four times: (1) with a 300-Hz context tone in both the prime and the probe presentation, (2) with a 700-Hz context tone in the prime and the probe presentation (these implementations will be referred to as context repeated trials), (3) with a 300-Hz context tone in the prime presentation and a 700-Hz context tone in the probe presentation, and (4) with a 700-Hz context tone in the prime presentation and a 300-Hz context tone in the probe presentation (there implementations will be referred to as context changed trials). This set of 192 trials was duplicated, resulting in 384 experimental trials, which were presented in a random sequence. For each of the 384 trials, it was randomly decided whether the attended prime was presented to the right or the left ear. The attended probe was always presented to the other ear.
Procedure
Participants were seated in front of the computer und adjusted the headphones. Then, they were introduced to the experimental sounds and the task. In a first training, they heard sound pair presentations that were preceded by the metronome click indicating the to-be-attended ear. Responses on the USB-connected response box were followed by audiovisual feedback about the correctness of each response. Participants entered the next training when 60% of the preceding 15 responses had been correct. If participants did not reach this criterion within 50 trials, they were given a choice to quit or to start again with the training. The second training was identical to the first with the only difference that context tones were added to the sound pair presentations. Participants were told that the context tones were irrelevant to the task and therefore should be ignored. After reaching the learning criterion of the second training, participants responded to six prime-probe trials using the timing of the subsequent experimental trials.
Before entering the experiment proper, participants were asked to be as accurate as possible while trying to respond fast. Each experimental trial began with the metronome click at the to-be-attended prime side, followed by a 500-ms cue-target interval and the prime presentation. After the prime response, a response-to-stimulus interval of 500 ms was followed by the metronome click that cued the to-be-attended probe side. The probe click was always presented to the side opposite to that of the prime. The click was followed by a 500-ms cue-target interval before the probe presentation was presented.
Participants were given feedback about the correctness of their response after each prime-probe trial. The feedback was followed by a 1,200-ms intertrial interval. Responses faster than 100 ms and slower than 3,000 ms were counted as invalid. After every block of 20 prime-probe trials, participants received a summary feedback about both their average reaction time and their error percentage. They started the next block at their own discretion. After the last experimental trial, participants were informed about the purpose of the experiment. Testing lasted approximately 45-60 minutes.
Design
The experiment comprised a 2 × 2 within-subject design with trial type (ignored repetition vs. control) and prime-probe context (repeated vs. changed) as independent variables. The dependent variables were participants’ average reaction time and error rates. To test whether the probability of prime response retrieval was increased for ignored repetition compared with control trials and whether this increase was modulated by prime-probe context, probe error frequencies were accumulated and analyzed across participants. To this end, the multinomial model described in Mayr and Buchner (2006) was used.Footnote 4 Furthermore, adjusted error rates were calculated by subtracting the prime response errors from the overall error rates for each participant and each experimental condition.
A difference in the size of the negative priming effect between trials with a prime-probe context repetition and a prime-probe context change was central to the hypothesis tested. To detect a medium difference in the size of the negative priming effect between context repeated and context changed trials (i.e. f = 0.25 as defined by Cohen, 1988), given desired levels of α = β = .05 and an assumed correlation of ρ = .4 between the negative priming effects in both context conditions, data had to be collected from a sample of 65 participants (Faul, Erdfelder, Lang, & Buchner, 2007). The final sample comprised 58 participants, so that the power was slightly smaller (.93) than what we had planned for. The significance level was set to .05 for all statistical tests. To prevent α error inflation the Bonferroni-Holm procedure (Holm, 1979) was applied whenever one-sided follow-up tests were needed to test whether the negative priming effect was statistically significant in the context repeated and the context changed condition.
Results
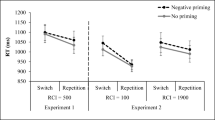
Probe responses were slower in ignored repetition than in control trials whereas the context manipulation did not affect response times (Fig. 1, upper panel). Accordingly, a two-factorial MANOVA of the probe reaction times with trial type (ignored repetition vs. control) and context (repeated vs. changed) as within-subject variables showed a main effect of trial type, F(1, 57) = 44.87, p < .01, ηp2 = .44, but neither the main effect of context, F(1, 57) = 0.12, p = .73, ηp2 < .01, nor the interaction was significant, F(1, 57) = 0.75, p = .39, ηp2 = .01.

Mean probe reaction times in Experiment 1 (upper panel) and Experiment 2 (lower panel) as a function of trial type (ignored repetition vs. control) and prime-probe context (repeated vs. changed). The error bars depict the standard errors of the means
Probe responding in ignored repetition trials comprised more errors than responding in control trials (Fig. 2, upper panel). This effect was larger when the context was repeated than when it was changed. A two-factorial MANOVA of the probe error rates with trial type (ignored repetition vs. control) and context (repeated vs. changed) as within-subject variables revealed a significant main effect of trial type, F(1, 57) = 20.15, p < .01, ηp2 = .26, and a significant interaction between trial type and context, F(1, 57) = 5.96, p = .02, ηp2 = .09, whereas the main effect of context was not significant, F(1, 57) = 2.04, p = .16, ηp2 = .03. Follow-up tests revealed that the negative priming effect in error rates was significant in the context repeated condition, t(57) = 4.42, p < .01, dz = 0.58, and in the context changed condition, t(57) = 2.31, p = .01, dz = 0.30, but the sample effect size was clearly smaller when the context changed than when it was repeated.

Probe error rates comprise all types of errors (response frequencies categorized by error type are presented in Table 2). To analyze whether the interaction between trial type and context was caused by an increase in prime response retrieval errors in the context repeated condition, we ran a multinomial analysis of the error data. The prime response retrieval parameters prrIR for the ignored repetition condition and prrC for the control condition are depicted in Fig. 3 (upper panel). Initially, we tested in each context condition the goodness-of-fit of the model with the restriction prrIR = prrC. This restricted model had to be rejected for the context repeated condition, G2(1) = 35.25, p < .01Footnote 5, ω = .08, whereas the misfit of the restricted model just missed the preset level of significance in the context changed condition, G2(1) = 3.20, p = .07, ω = .02. This implies that there was clear evidence of prime response retrieval processes in the context repeated condition but not in the context changed condition.

Probability estimates for the model parameters representing the probability of prime response retrieval (prr) in Experiment 1 (upper panel) and Experiment 2 (lower panel) as a function of trial type (ignored repetition vs. control) and prime-probe context (repeated vs. changed). The error bars depict the standard errors of the means
We then compared the prr parameters across context conditions by combining the models for the two context conditions into one processing tree. In a first step, the prrC parameters were set to be equal across context conditions. The restricted model fit the data, G2(1) = 0.71, p = .40, ω = .01, implying that a pure context repetition did not increase prime response retrieval processes. In a second step, the prrIR parameters also were set to be equal across context conditions. The nested model with this additional restriction did not fit the data, G2(1) = 9.06, p < .01, ω = .03, implying that the prime response retrieval process induced by the repetition of the distractor stimulus was stronger in the context repeated than in the context changed condition.
The results of the multinomial analysis were corroborated by an additional error analysis in which we analyzed the adjusted error rates (overall errors without the prime response errors). There was a significant main effect of trial type, F(1, 57) = 4.45, p = .04, ηp2 = .07, but neither an effect of context, F(1, 57) = 0.20, p = .66, ηp2 < .01, nor a significant interaction between both variables, F(1, 57) = 1.25, p = .27, ηp2 = .02. This implies that the negative priming effect in the adjusted errors did not differ between context repeated and context changed trials.
Experiment 2
Method
Participants
Participants were 61 adults (19 males), most of whom were University of Passau students. Their age ranged from 18 to 33 years (M = 22). Participants received monetary compensation for their participation. All participants reported normal hearing. Data sets of three further participants had to be excluded. They committed more than 50% erroneous responses in all hypothesis-relevant experimental conditions (compared with an average of approximately 9% in all relevant experimental conditions), which strongly suggests either a misunderstanding of the task or a lack of willingness to follow the instructions.
Materials, task, and procedure
Materials, task, and procedure were identical to those of Experiment 1 with the following exceptions. The sounds were played over headphones that were plugged into a Windows computer. The Windows version of the experimental software (LiveCode) that controlled stimulus presentation in Experiment 1 was used in Experiment 2. As a result of pretesting, the sounds were set to an easily audible and comfortable loudness comparable to Experiment 1, but loudness level was not measured.
Response buttons were the sagittally aligned “9,” “6,” “3,” and “,” keys on the number pad of an USB-connected Cherry MX Board 6.0 keyboard. A total of 31 participants had to press the two distal buttons (frog and piano) with the middle and index fingers of their right hands and the two proximal buttons (drum and bell) with the middle and index fingers of their left hands. For the remaining 30 participants, arrangement was reversed.
Design
The design of Experiment 2 was identical to that of Experiment 1. To detect a medium difference in the size of the negative priming effect between context repeated and context changed trials (i.e., f = 0.25 as defined by Cohen, 1988), given desired levels of α = β = .05 and an assumed correlation of ρ = .4 between the negative priming effects in both context conditions, data had to be collected from a sample of 65 participants (Faul et al., 2007). The final sample comprised 61 participants, so that the power was slightly smaller (.94) than what we had planned for.
Results
As expected, probe responses were slower in ignored repetition than in control trials. In contrast to Experiment 1, responses were slightly slowed down in trials with a context repetition (Fig. 1, lower panel), but as in Experiment 1, there was no difference in the size of the negative priming effect for trials with a context change and a context repetition. A two-factorial MANOVA of the probe reaction times with trial type (ignored repetition vs. control) and context (repeated vs. changed) as within-subject variables showed a main effect of trial type, F(1, 60) = 78.76, p < .01, ηp2 = .57, a main effect of context, F(1, 60) = 6.34, p = .01, ηp2 = .10, but the interaction between both variables was not significant, F(1, 60) = 0.17, p = .69, ηp2 < .01.
As in Experiment 1, there was a negative priming effect in probe error rates, which was larger when the context repeated than when it changed (Fig. 2, lower panel). The descriptive pattern of results was reflected in the statistical analysis. A two-factorial MANOVA of the probe error rates with trial type (ignored repetition vs. control) and context (repeated vs. changed) as within-subject variables revealed a significant main effect of trial type, F(1, 60) = 29.18, p < .01, ηp2 = .33, and a significant interaction between trial type and context, F(1, 60) = 10.63, p < .01, ηp2 = .15, whereas the main effect of context was not significant, F(1, 60) = 0.22, p = .64, ηp2 < .01. Follow-up tests revealed that the negative priming effect in error rates was significant in the context repeated condition, t(60) = 6.36, p < .01, dz = 0.81, and in the context changed condition, t(60) = 2.48, p = .01, dz = 0.32, but the sample effect size was clearly smaller when the context changed than when it was repeated.
The response frequencies categorized by error type are presented in Table 2. The prime response retrieval parameters prrIR and prrC are depicted in Fig. 3 (lower panel). We tested in each context condition the goodness-of-fit of the model with the restriction prrIR = prrC. This restricted model had to be rejected for the context repeated condition, G2(1) = 39.76, p < .01, ω = .09, whereas the misfit of the restricted model was just slightly below the preset criterion for statistical significance in the context changed condition, G2(1) = 4.00, p = .05 (.046), ω = .03. This implies that there was clear evidence of prime response retrieval processes in the context repeated condition and in the context changed condition, but the effect was much larger in the context repeated condition than in the context changed condition.
As in Experiment 1, we then compared the prr parameters across context conditions by combining the models for the two context conditions into one processing tree. In a first step, the prrC parameters were set to be equal across context conditions. The restricted model fit the data, G2(1) = 0.62, p = .43, ω < .01, implying that a pure context repetition did not increase prime response retrieval processes. In a second step, the prrIR parameters were also set to be equal across context conditions. The nested model with the additional restriction did not fit the data, G2(1) = 13.57, p < .01, ω = .04, implying that the prime response retrieval process was stronger in the context repeated than in the context changed condition.
Again, the results of the multinomial analysis were corroborated by an additional error analysis in which the adjusted error rates (overall errors without the prime response errors) were analyzed. There was a significant main effect of trial type, F(1, 60) = 8.23, p = .01, ηp2 = .12, but neither an effect of context, F(1, 60) = 1.24, p = .27, ηp2 = .02, nor a significant interaction between both variables, F(1, 60) = 2.22, p = .14, ηp2 = .04. This implies that there was no difference in the negative priming effect in error rates left when errors due to prime response retrieval had been excluded.
General Discussion
The present experiments were planned to test whether contextual similarity influences the negative priming effect (i.e., the difference between ignored repetition and control trials) in the auditory modality and to analyze the processes underlying the possible modulation by context. The findings of Experiment 1 and their replication in Experiment 2 consistently show a larger negative priming effect for high context similarity (context repeated trials) than for low context similarity (context changed trials). However, the increase in negative priming with context repetition was only found in the error data. The negative priming effect in response times was insensitive to contextual similarity in both experiments.
An analysis of the response frequencies using a multinomial measurement model showed that prime response retrieval was substantial in the context repeated condition whereas this was much less so in the context changed condition. In other words, it seems that contextual similarity between prime and probe facilitates episodic retrieval processes, rendering the retrieval of prime response information more likely. Most interestingly, the prime response retrieval mechanism seems to be the sole mechanism involved in negative priming that is sensitive to contextual similarity. This can be inferred from the finding that there was neither a context modulation of negative priming effects in the reaction times nor any evidence of contextual effects in the adjusted error analysis after removing the prime response errors.Footnote 6
However, the fact that a negative priming effect is still evident in the error rates after removing the prime response errors indicates that prime response retrieval cannot be the only process involved in the auditory negative priming effect. Other processes—such the retrieval of nonresponse information (Neill & Valdes, 1992) or the inhibition of distractor representations (Houghton & Tipper, 1994; Tipper, 1985)—have to be assumed to account for the remaining negative priming effects in the error rates as well as in the reaction times.
Past research has found several differences in negative priming, and the processes contributing to the effect between modalities. For example, auditory negative priming seems to have a stronger automatic component und is less strategically modifiable than visual negative priming (Frings & Wentura, 2008; Mayr & Buchner, 2010b). The present findings, in contrast, reveal a commonality between modalities in showing for the first time that negative priming in the auditory modality can be modulated by contextual similarity, similar to what has been reported for the visual modality (Chao, 2009; Chao & Yeh, 2008; Neill, 1997). However, such a commonality at the level of the measured negative priming effects does not necessarily imply that the underlying mechanisms are the same. We found a contextual modulation of auditory negative priming that was exclusively based on prime response retrieval processes. It is not clear whether the same is true for the visual modality. To our knowledge, the study by Tse et al. (2011) is the only study in which the processes underlying the contextual modulation of visual negative priming were examined. Based on analyses of the reaction time distributions, Tse et al. (2011, Experiment 2) found that the negative priming effect increased with increasing reaction times only for same context trials but not for different context trials. This finding was interpreted as evidence of episodic retrieval processes in the same context condition because a slower processing of the probe targets (as indicated by increasing reaction times) makes retrieving prime information incompatible with probe responding more likely. This should lead to a larger negative priming effect. However, the design of the experiment by Tse et al. (2011) was not suitable to differentiate between prime response retrieval and original episodic retrieval (nonresponse retrieval) accounts. To answer the question whether prime response retrieval is the driving force in the visual modality, an experiment manipulating context repetition in a visual variant of the four-alternative forced choice reaction time task should be the next step.
As mentioned in the Introduction, the present findings can be interpreted against the background of the event-file framework (Hommel, 1998). The increase in errors with the prime response when the prime distractor is repeated as the probe target (i.e., in Ignored Repetition trials) suggests that prime distractor and prime response were bound together during prime processing into an event file. This event file is supposed to be reactivated by reencountering one of its components. In the present study, the likelihood of retrieving prime response information cued by the probe target (when it was a repetition of the prime distractor) was increased when the context was repeated. This suggests that contextual information also was included in the event file and acted as an additional retrieval cue. However, context repetition alone—as it occurred in the context repeated control trials—did not increase the probability of prime response retrieval. This can be inferred from the fact that the prrc parameters did not differ between the context repeated and the context changed control trials. It seems as if the repetition of the task-irrelevant context sound per se is a weak or even ineffective direct cue to the prime response whereas it can boost the effect of the repeated distractor stimulus on retrieving the prime response. Such a catalyst effect of the context cue does not have a straightforward interpretation within the event file framework which does not allow for such asymmetrically effective retrieval cues without additional assumptions. Specifically, the asymmetrical effect of context observed here is incompatible with the standard assumption that simple binary bindings (i.e., assuming that always two features are tied together) in the event file are established during encoding (Hommel, 2007). One such binding should be that between the prime distractor and the prime response and another one should be that between the prime context and the prime response. The reinstatement of the prime distractor as the target in the probe presentation and the reinstatement of the prime context in the probe presentation should independently serve as cues to the prime episode and, hence, the prime response. From the independence it follows that the effects of those two cues should be additive, which was obviously not the case. Reinstating the prime context affected prime response retrieval only when the prime distractor was reinstated as well.Footnote 7
In contrast to the present findings, previous research using the distractor-response binding task (Frings, Rothermund, & Wentura, 2007) suggested that stimulus-response bindings can be retrieved independently (Giesen & Rothermund, 2014). The reasons for this difference between previous research and our present findings are not yet understood and will have to be clarified in future research. For instance, in the present study the context was relatively simple, not very variable, and consistently task-irrelevant. Also, targets, distractors, and context stimuli were auditory rather than visual. Finally, procedural differences between negative priming and the distractor-response binding task (Frings et al., 2007) may account for the difference.
Prospective studies in the auditory modality need to analyze the principles of auditory context integration and retrieval. For example, it would be interesting to know whether the integration of auditory context depends on sound properties of the context stimulus and follows analog perceptual principals (such as figure-ground segmentation) as is the case in the visual modality (Frings & Rothermund, 2017). For example, it is conceivable that the simple and monotone context sounds were automatically perceived as backgrounds behind the more distinct and variable distractor sounds (i.e., figures). While figure-like stimuli might function as individual retrieval cues, background sounds might only be perceived as part of the whole scenario (including the foreground), and hence, are only effective retrieval cues in this foreground-background compound. Possibly, factors that influence auditory grouping such as similarity in timbre or pitch or proximity in space (Bregman, 1993) might be crucial for finding after-effects of binding between context, distractor, and responses. Furthermore, variables, such as stimulus salience, task relevance, and temporal overlap (between stimuli and responses), that have been shown to be crucial dimensions for the binding of event files (Hommel, 2004; Zmigrod & Hommel, 2009) also should be analyzed in their importance for context integration and retrieval.
Independent of these unanswered questions, the results of the present findings are quite clear-cut in showing for the first time that auditory negative priming is modulated by prime-probe contextual similarity as predicted by episodic retrieval accounts. What is more, the analysis of the error data has shown that prime-probe contextual similarity boosts prime response retrieval processes selectively.
Change history
05 September 2018
The Publisher regrets that two erroneous values were introduced by the typesetter when performing proof corrections.
Notes
These authors ran distributional analyses of reaction times to examine the mechanisms underlying the negative priming effect. Details of their study will be described in the General Discussion section.
Note that this testing rationale is not applicable to the negative priming effect in reaction times. However, to anticipate, negative priming in reaction times was not at all modulated by contextual similarity.
Following the described algorithm to generate experimental trials, 24 different ignored repetition as well as 24 different attended repetition trials can be constructed. However, for this entire set of trials, there is always one ignored repetition trial and one attended repetition trial that share the same control trial. Using the entire set of trials, every control trial would occur twice as often as every ignored repetition or attended repetition trial. To avoid this confound, ignored repetition and attended repetition trials were systematically assigned to Set 1 or Set 2. To this end, identical control trials were assigned to different sets. Sets 1 and 2 were completely parallel with respect to the occurrences of the different tones as well as the occurrences of the combinations of attended and ignored tones. In each set, the prime target and response did not predict the probe target and response (for a more detailed description of the two sets and their properties, see Mayr and Buchner 2006). Participants were randomly assigned to Set 1 or 2.
The multinomial model described in Mayr and Buchner (2006) represents the cognitive processes that are presumably involved in probe responding in a four alternative forced choice response task. According to the model, correct identifications and, as a consequence, correct responses to the probe target take place with probability ci. In contrast, erroneous responding might be the result of different processes. Errors might be caused by probe stimulus confusions, that is, by inadvertently confusing target and distractor stimuli, resulting in a response with the probe distractor (with probability psc). If probe stimulus confusion is not dominating responding, prime response retrieval (prr) processes might result in probe errors with the former prime response. Alternatively, an erroneous response with the forth response option can be given. Multinomial modeling does not only require to specify the assumed underlying processes, it also allows to test specific predictions concerning the model parameters. For example, the prime response retrieval model predicts that the conditional probability of prime response retrieval should be larger in the Ignored Repetition trials than in the Control trials (prrIR > prrc ). To test this prediction, the goodness-of-fit of the model with the restriction prrIR = prrc will be tested. A significant misfit of this model is to be expected when the prime response retrieval account is valid.
All multinomial processing tree analyses were run with multiTree (Moshagen, 2010).
A closer look at the categorical response data in Table 2 reveals that the frequency of probe distractor errors was somewhat larger in Ignored Repetition trials than in Control trials and that this increase was stronger in context repeated than in context changed trials. Possibly, participants tagged prime distractor stimuli with “do-not-respond” flags (consistent with the initially proposed episodic retrieval account by Neill et al., 1992). Retrieving this information in the probe of Ignored Repetition trials due to the repetition of the prime distractor stimulus would make it harder to execute the correct probe response, allowing for occasional intrusions of the probe distractor. The fact that the increase in probe distractor errors was stronger for context repeated than for context changed trials suggests that context might have functioned as an additional retrieval cue to the nonresponse information attached to the prime distractor. We thank Trammell Neill for suggesting this additional mechanism prone to prime-probe contextual similarity. However, this potential second mechanism seems to be a rather weak one which was not reliably measurable in the present set-up as our additional error analyses of the adjusted error rates (overall errors without the prime response errors) revealed: In both experiments, there was neither a significant main effect of context nor a significant interaction between context and trial type after the prime response errors had been eliminated
Whereas a context repetition was only effective in retrieving the prime response when the distractor stimulus was repeated, too, the reversed relation was not observed: Solely repeating the distractor (but not the context) led to an increase in prime response errors. This implies that prime response retrieval could either take place based on distractor repetitions or based on the repetition of the compound of distractor and context. Possibly, differences in sound quality and/or in task relevance might be responsible for the found differences in retrieval effectiveness of context and distractor stimuli (see further below in the text). However, the present study was neither intended nor able to differentiate between these possibilities.
References
Bregman, A. S. (1993). Auditory scene analysis: Hearing in complex environments. Thinking in sound: The cognitive psychology of human audition (pp. 10–36). New York, NY: Clarendon Press/Oxford University Press; US.
Chao, H.-F. (2009). Revisiting the prime-probe contextual similarity effect on negative priming: The impact of cue variability. European Journal of Cognitive Psychology, 21(4), 484–500. https://doi.org/10.1080/09541440802049051
Chao, H.-F., & Yeh, Y. Y. (2008). Attentional demand and memory retrieval in negative priming. Psychological Research/Psychologische Forschung, 72(3), 249–260. https://doi.org/10.1007/s00426-006-0106-y
Cohen, J. (1988). Statistical power analysis for the behavioral sciences (2nd). Hillsdale: Lawrence Erlbaum Associates.
D’Angelo, M. C., Thomson, D. R., Tipper, S. P., & Milliken, B. (2016). Negative priming 1985 to 2015: a measure of inhibition, the emergence of alternative accounts, and the multiple process challenge. Quarterly Journal of Experimental Psychology, 69(10), 1890–1909. https://doi.org/10.1080/17470218.2016.1173077
Faul, F., Erdfelder, E., Lang, A.-G., & Buchner, A. (2007). G*Power 3: a flexible statistical power analysis program for the social, behavioral, and biomedical sciences. Behavior research methods, 39(2), 175–191. https://doi.org/10.3758/BF03193146
Fox, E., & de Fockert, J. W. (1998). Negative priming depends on prime-probe similarity: Evidence for episodic retrieval. Psychonomic Bulletin & Review, 5(1), 107–113. https://doi.org/10.3758/BF03209464
Frings, C., Koch, I., & Moeller, B. (2017). How the mind shapes action: Offline contexts modulate involuntary episodic retrieval. Attention, Perception, & Psychophysics, 79(8), 2449–2459. https://doi.org/10.3758/s13414-017-1406-6
Frings, C., & Rothermund, K. (2017). How Perception Guides Action: Figure-Ground Segmentation Modulates Integration of Context Features into S-R Episodes. Journal of Experimental Psychology: Learning, Memory, and Cognition. https://doi.org/10.1037/xlm0000403
Frings, C., Rothermund, K., & Wentura, D. (2007). Distractor repetitions retrieve previous responses to targets. Quarterly Journal of Experimental Psychology, 60, 367–1377. https://doi.org/10.1080/17470210600955645
Frings, C., Schneider, K. K., & Fox, E. (2015). The negative priming paradigm: An update and implications for selective attention. Psychonomic Bulletin & Review, 22(6), 1577–1597. https://doi.org/10.3758/s13423-015-0841-4
Frings, C., & Wentura, D. (2008). Separating context and trial-by-trial effects in the negative priming paradigm. European Journal of Cognitive Psychology, 20(2), 195–210. https://doi.org/10.1080/17470910701363090
Giesen, C., & Rothermund, K. (2014). Distractor repetitions retrieve previous responses and previous targets: experimental dissociations of distractor-response and distractor-target bindings. Journal of Experimental Psychology-Learning Memory and Cognition, 40(3), 645–659. https://doi.org/10.1037/a0035278
Holm, S. (1979). A simple sequentially rejective multiple test procedure. Scandinavian Journal of Statistics, 6, 65–70.
Hommel, B. (1998). Event files: Evidence for automatic integration of stimulus-response episodes. Visual Cognition, 5(1-2), 183–216. https://doi.org/10.1080/713756773
Hommel, B. (2004). Event files: Feature binding in and across perception and action. Trends in Cognitive Sciences, 8(11), 494–500. https://doi.org/10.1016/j.tics.2004.08.007
Hommel, B. (2007). Feature integration across perception and action: Event files affect response choice. Psychological Research, 71, 43–63. https://doi.org/10.1007/s00426-005-0035-1
Houghton, G., & Tipper, S. P. (1994). A model of inhibitory mechanisms in selective attention. In D. Dagenbach & T. H. Carr (Eds.), Inhibitory mechanisms of attention, memory, and language (pp. 53–112). San Diego: Academic Press.
Logan, G. D. (1988). Toward an instance theory of automatization. Psychological Review, 95(4), 492–527.
Mayr, S., & Buchner, A. (2006). Evidence for episodic retrieval of inadequate prime responses in auditory negative priming. Journal of Experimental Psychology: Human Perception and Performance, 32(4), 932–943. https://doi.org/10.1037/0096-1523.32.4.932
Mayr, S., & Buchner, A. (2010a). Auditory negative priming endures response modality change; prime-response retrieval does not. Quarterly Journal of Experimental Psychology, 63(4), 653–665. https://doi.org/10.1080/17470210903067643
Mayr, S., & Buchner, A. (2010b). Episodic retrieval processes take place automatically in auditory negative priming. European Journal of Cognitive Psychology, 22(8), 1192–1221. https://doi.org/10.1080/09541440903409808
Mayr, S., & Buchner, A. (2014). Intact episodic retrieval in older adults: Evidence from an auditory negative priming task. Experimental Aging Research, 40(1), 13–39. https://doi.org/10.1080/0361073X.2014.857541
Mayr, S., Buchner, A., & Dentale, S. (2009a). Prime retrieval of motor responses in negative priming. Journal of Experimental Psychology: Human Perception and Performance, 35(2), 408–423. https://doi.org/10.1037/0096-1523.35.2.408
Mayr, S., Buchner, A., Möller, M., & Hauke, R. (2011a). Spatial and identity negative priming in audition: Evidence of feature binding in auditory spatial memory. Attention, Perception, & Psychophysics, 73(6), 1710–1732. https://doi.org/10.3758/s13414-011-0138-2
Mayr, S., Hauke, R., & Buchner, A. (2009b). Auditory location negative priming: A case of feature mismatch. Psychonomic Bulletin & Review, 16(5), 845–849. https://doi.org/10.3758/PBR.16.5.845
Mayr, S., Möller, M., & Buchner, A. (2011b). Evidence of vocal and manual event files in auditory negative priming. Experimental Psychology, 58(5), 353–360. https://doi.org/10.1027/1618-3169/a000102
Milliken, B., Joordens, S., Merikle, P. M., & Seiffert, A. E. (1998). Selective attention: A reevaluation of the implications of negative priming. Psychological Review, 105(2), 203–229. https://doi.org/10.1037/0033-295X.105.2.203
Moore, C. M. (1996). Does negative priming imply preselective identification of irrelevant stimuli? Psychonomic Bulletin & Review, 3(1), 91–94. https://doi.org/10.3758/BF03210746
Moshagen, M. (2010). multiTree: A computer program for the analysis of multinomial processing tree models. Behavior Research Methods, 42(1), 42–54. https://doi.org/10.3758/BRM.42.1.42
Neill, W. T. (1997). Episodic retrieval in negative priming and repetition priming. Journal of Experimental Psychology: Learning, Memory, and Cognition, 23(6), 1291–3105. https://doi.org/10.1037/0278-7393.23.6.1291
Neill, W. T., & Valdes, L. A. (1992). Persistence of negative priming: Steady state or decay? Journal of Experimental Psychology: Learning, Memory, and Cognition, 18(3), 565–576. https://doi.org/10.1037/0278-7393.18.3.565
Neill, W. T., Valdes, L. A., Terry, K. M., & Gorfein, D. S. (1992). Persistence of negative priming: II. Evidence for episodic trace retrieval. Journal of Experimental Psychology: Learning, Memory, and Cognition, 18(5), 993–1000. https://doi.org/10.1037/0278-7393.18.5.993
Park, J., & Kanwisher, N. (1994). Negative priming for spatial locations: Identity mismatching, not distractor inhibition. Journal of Experimental Psychology: Human Perception and Performance, 20(3), 613–623. https://doi.org/10.1037/0096-1523.20.3.613
Pashler, H. E., & Baylis, G. C. (1991). Procedural learning: II. Intertrial repetition effects in speeded-choice tasks. Journal of Experimental Psychology: Learning, Memory, and Cognition, 17(1), 33-48.
Rothermund, K., Wentura, D., & De Houwer, J. (2005). Retrieval of incidental stimulus-response associations as a source of negative priming. Journal of Experimental Psychology: Learning, Memory, and Cognition, 31(3), 482–495. https://doi.org/10.1037/0278-7393.31.3.482
Smith, S. M., & Vela, E. (2001). Environmental context-dependent memory: A review and meta-analysis. Psychonomic Bulletin & Review, 8(2), 203–220. https://doi.org/10.3758/bf03196157
Stolz, J. A., & Neely, J. H. (2001). Taking a bright view of negative priming in the light of dim stimuli: Further evidence for memory confusion during episodic retrieval. Canadian Journal of Experimental Psychology, 55(3), 219–230. https://doi.org/10.1037/h0087368
Tipper, S. P. (1985). The negative priming effect: Inhibitory priming by ignored objects. Quarterly Journal of Experimental Psychology: Human Experimental Psychology, 37A(4), 571–590. https://doi.org/10.1080/14640748508400920
Tse, C.-S., Hutchison, K. A., & Li, Y. (2011). Effects of contextual similarity and target-repetition proportion on negative priming in RT distributional analyses. Journal of Experimental Psychology: Human Perception and Performance, 37(1), 180–192. https://doi.org/10.1037/a0019930
Tulving, E. (1983). Elements of episodic memory. Oxford: Oxford University Press.
Wong, K. F. E. (2000). Dissociative prime-probe contextual similarity effects on negative priming and repetition priming: A challenge to episodic retrieval as a unified account of negative priming. Journal of Experimental Psychology: Learning, Memory, and Cognition, 26(6), 1411–1422. https://doi.org/10.1037/0278-7393.26.6.1411
Zmigrod, S., & Hommel, B. (2009). Auditory event files: integrating auditory perception and action planning. Attention, Perception, & Psychophysics, 71(2), 352–362. https://doi.org/10.3758/APP.71.2.352
Author information
Authors and Affiliations
Corresponding author
Additional information
The original version of this article was revised: The Publisher regrets that two erroneous values were introduced by the typesetter when performing proof corrections. The first error occurred in the “Design” section of Experiment 1 near the end of the second paragraph: “ρ = .04” should be “ρ = .4”
The second error occurred at the very end of the “Experiment 1” section in the “Appendix”: “dz = .010” should be “dz = 0.10”
Appendix
Appendix
Analysis of the Attended Repetition trials
Attended Repetition and Attended Repetition Control trials were included in Experiment 1 and 2 in order to avoid stimulus and response contingencies between prime and probe that could have resulted in strategic responding. However, from the perspective of the encoding specificity hypothesis (Tulving, 1983), repetition priming effects due to prime-to-probe target and response repetitions also should be moderated by prime-to-probe contextual similarity (as shown in the visual modality, see Neill, 1997). We will briefly summarize the results of the Attended Repetition trials and their controls and discuss the findings against the background of the literature.
Experiment 1
Probe responses were faster in attended repetition than in attended repetition control trials, whereas the context manipulation did not affect response times (Fig. 4, upper panel). Accordingly, a two-factorial MANOVA of the probe reaction times with trial type (attended repetition vs. attended repetition control) and context (repeated vs. changed) as within-subject variables showed a main effect of trial type, F(1, 57) = 19.76, p < .01, ηp2 = .26, but neither the main effect of context, F(1, 57) = 2.70, p = .11, ηp2 = .05, nor the interaction was significant, F(1, 57) = 2.29, p = .14, ηp2 = .04.

Concerning probe accuracy, there was a slight decrease in response errors in the attended repetition trials as compared with attended repetition control trials, but this effect was only evident in the context repeated trials (Fig. 5, upper panel). A two-factorial MANOVA of the probe error rates with trial type (attended repetition vs. attended repetition control) and context (repeated vs. changed) as within-subject variables revealed neither a significant main effect of trial type, F(1, 57) = 0.41, p = .52, ηp2 = .01, nor a significant main effect of context, F(1, 57) = 2.13, p = .15, ηp2 = .04. However, the interaction between trial type and context exceeded the criterion of significance, F(1, 57) = 4.49, p = .04, ηp2 = .07. Follow-up tests (one-sided) revealed that there was a positive priming effect in error rates in the context repeated condition, t(57) = −2.03, p = .02, dz = 0.27, but not in the context changed condition, t(57) = 0.75, p = .23, dz = 0.10.

Experiment 2
Probe responses were faster in attended repetition than in attended repetition control trials and context repetition led to faster responses than context change. Furthermore, the reaction time decrease due to repetition of the prime target was larger in the same context condition than in the different context condition (Fig. 4, lower panel). A two-factorial MANOVA of the probe reaction times with trial type (attended repetition vs. attended repetition control) and context (repeated vs. changed) as within-subject variables revealed a main effect of trial type, F(1, 60) = 44.66, p < .01, ηp2 = .43, a main effect of context, F(1, 60) = 17.88, p < .01, ηp2 = .23, as well as an interaction between both variables, F(1, 60) = 19.13, p < .01, ηp2 = .24. Follow-up tests (one-sided) revealed that the decrease in reaction times in the attended repetition trials was significant in both context conditions, but clearly larger in the context repeated condition, t(60) = −7.50, p < .01, dz = 0.96, than in the context changed condition, t(60) = −3.45, p < .01, dz = 0.44.
Concerning probe accuracy, there was a slight decrease in response errors in trials with a context repetition compared with trials with a context change, but the manipulation of trial type did not seem to have an effect (Fig. 5, lower panel). A two-factorial MANOVA of the probe error rates with trial type (attended repetition vs. attended repetition control) and context (repeated vs. changed) as within-subject variables revealed neither a main effect of trial type, F(1, 60) < .01, p = .96, ηp2 < .01, nor a significant interaction between trial type and context, F(1, 60) = 3.37, p = .07, ηp2 = .05. However, the main effect of context was significant, F(1, 60) = 7.07, p = .01, ηp2 = .11.
Discussion
Both experiments revealed faster responding in attended repetition trials with a prime-to-probe stimulus and response repetition as compared with the respective control trials without any stimulus and response repetitions. Such repetition effects are most likely caused by directly linking early stimulus representations and specific responses thereby short-cutting the response selection stage (Pashler & Baylis, 1991). Note that this explanation is also compatible with the idea that individual occurrences of stimuli and responses are bound into common episodes (or event files) that are automatically retrieved when parts (or all) of its elements are repeated (Hommel, 1998). In the case of attended repetition trials, the retrieved episode would be completely identical to the current situation, which would facilitate responding.
Furthermore, in both experiments, context repetitions between prime and probe led to an increase in repetition priming effects. In accordance with the present findings but for the visual modality, Neill (1997) demonstrated that repetition priming effects like negative priming effects were modulated by prime-to-probe contextual similarity. On the assumption that repetition priming effects are based on an episodic retrieval process, the present finding is to be expected from the perspective of the encoding specificity hypothesis (Tulving, 1983). According to this hypothesis, the retrieval of previous processing episodes depends on the similarity between encoding (prime) and retrieval (probe) context.
Although the core findings in both experiments were equivalent, Experiment 1 and Experiment 2 differed in the exact way context affected responding: The modulating effect of context was present in the error data in Experiment 1 but in the reaction times of Experiment 2. Furthermore, the modulating effect of context was larger in Experiment 2 (ηp2 = .24) than in Experiment 1 (ηp2 = .07). Note that the overall reaction time level was a little bit slower in Experiment 2 than in Experiment 1 (Figs. 1 and 4). Hence, there was somewhat more room for improvement (i.e., speed-up) in Experiment 2, which might be the reason why the modulation by context was found in the reaction times (and not in the error rates) in this experiment. Regardless of these experimental peculiarities, both experiments revealed the same crucial finding, that is, the dependence of repetition priming effects on contextual similarity between prime and probe.
Rights and permissions
About this article

Cite this article
Mayr, S., Möller, M. & Buchner, A. Contextual modulation of prime response retrieval processes: Evidence from auditory negative priming. Atten Percept Psychophys 80, 1918–1931 (2018). https://doi.org/10.3758/s13414-018-1574-z
Published:
Issue Date:
DOI: https://doi.org/10.3758/s13414-018-1574-z